(2023年1月16日更新) [ 日本語 | English ]
HOME > 講義・実習・演習一覧 / 研究概要 > 小辞典 > 生物学 > 藻類
藻類学 (phycology or alegology)藻類について科学研究する学問 |
海藻 = 海洋生態系の生産者 → その生態系を理解する上で重要 海藻 (かいそう) seeweed: 海に生息する藻類 海草 (かいそう、うみくさ) seegrass: 海に生息する維管束植物 研究者
|
|
葉状体: 多細胞海藻体構造
分裂組織
細胞壁 (cell wall)紅・褐・緑藻: 厚さ1 μm = クチクラ(≠ 陸上植物相同) (Hanic & Craigie 1969)構成成分: 炭水化物 (糸状(= 微小繊維)と不定形の2形状)・タンパク質 微小繊維: セルロース主。直径3-20 nm → セルロース: マンノースやキシロースの重合体 1974 Mackie & Preston: 微小繊維配列様式から細胞壁を2種類に分類バロニア型(交叉糸状型) crossed-fibrillar: 緑藻数種のみ。層状構造をした特異的なもの 繊維互いに角度90度で2方向に配列 多網型 multi-net construction: 他種。2つの部分からなる細胞壁持つ |
石灰化多くは紅藻。緑藻でもサボテングサ Halimeda 等が行なう
紅藻: 炭酸石灰が細胞壁基質中に蓄積され石灰化 鞭毛緑藻・褐藻の生殖細胞において見られる。紅藻にないa) 等毛型: 多くの緑藻 b) 不等毛型: 褐藻 c) 冠毛: 緑藻 Derbesia, Bryopsis特有 微小管細胞成長・分化に関与 (Green 1969)
微小管配列方向が最小繊維沈着様式を制御する → 最小繊維配列方向が細胞の形を制御する 生殖 reproduction
|
|
緑藻植物特徴形質 ≈ 車軸藻植物 → (Division Chlorophyta所属見解)。現世6属250種 外側に石灰質沈着し堅くなるものが多く化石として得られる
淡水生(湖多)固着生[主] + 汽水生。海産生未確認 形態主軸 main axis (ma): 茎状部 axis = 節細胞 node cell + 節間細胞 internode cell (in)
節間細胞: 1細胞からなる
輪生枝 vertical branch(let) or whorl (vb): 節部から周りへ放射状に伸びる 増殖栄養生殖: 球芽 bulbilによる無性生殖をするものあるが、遊走子見られない |
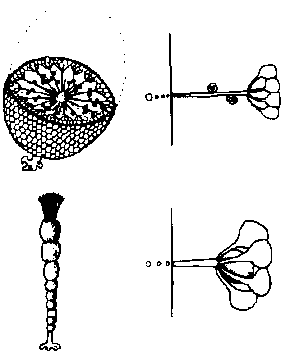
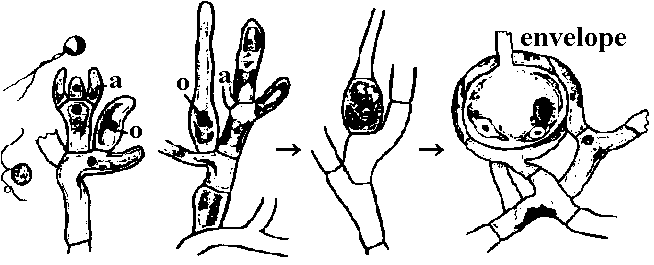
有性生殖: 普通、卵胞子oosporeと精子による卵生殖。特異的で複雑な構造の生殖器官有し造精器は、中空球形、壁は8枚の楯形細胞shield cellが接着構成される。楯形細胞内側中央にある柄細胞manubrial cell先端に造精糸antheridial filamentが房状につく。造精糸は精子母細胞antheridial mother cellが連接したもので各細胞内にS字状等長2本の尾型鞭毛持つ精子形成する。生卵器は多細胞からなる楕円状体で内部に1個の卵胞子があり、その周囲は螺旋状に捻れた壁細胞spiral cell層で包まれる。壁細胞先端には1-2個の小細胞が付着し、それらが小冠coronulaを形成する 造精器antheridium: 楯形細胞が8個周囲を囲まれ楯形細胞各々の内壁中央から内部に向かい把手細胞がでて、先端に球状細胞globoid cellつける。これから単列に連なる糸antheridial filamentが出て各細胞が精子になる 造卵器oogonium: 原糸体protonema(pr)の一方は仮板となる 卵胞子oosporeによる卵生殖 分類基準
等 |
|
分化した細胞器官持つ 同化色素: Chlorophyll a, b、カロチノイド(α, β-カロチン、アスタキサンチン、ルテイン、ネオキサンチン)等 葉緑体: 多重チラコイドラメラかグラナラメラ 貯蔵物質: デンプン 細胞壁主要構成物質: セルロース 単細胞 unicellular form、群体 colonial form 遊走細胞は一般に等長の鞭毛を前端に持つ。鞭毛はムチ型構造 体制進化段階 →
運動性鞭毛: 運動性なし non-mobile (Chlorococcales)↔ 運動性あり mobile (Volvocales) (s: 胞子体, G: 配偶体, z: 遊走子, g: 配偶子) 生活環 life cycle1) Spirogyra type アオミドロ型
単相のみ
__┏━━> ○ z ━━━━> ██████ G
__┏━━> ○ z ━━━━> ██████ G
__┏━━> ○ ━━━━━━━> ███ G
━_┏━━━━━━━━━━━━━┓ 系統と分類色素体微細構造、光合成色素、光合成産物、鞭毛構造、細胞壁物質等 → 陸生緑色植物と関係藻体上では非常に分化し緑藻植物の陸上への移行考えられる
体制進化を第一義的基準 → 次に細胞分裂様式(= 隔壁形成様式及び生殖方法を基準) 観察
Class Chlorophyceae 緑藻鋼単核Order Volvocales オオヒゲマワリ多くは淡水産藻体: 運動性単細胞or定数群体。2 or 4本の等長鞭毛 色素体: 杯状。1-数個ピレノイド 無性生殖: 2分裂 or 娘定数群体形成。同型又は異型配偶子の接合や卵生殖 Chlamydomonadaceae: Carteria, Chlamydomonas, Eudorina, Gonium, Pandorina, Pascheriella, Platidorina, Platymonas, Pleodorina, Volvox
配偶子は2本の鞭毛。異型配偶子接合 anisogamy Clamydomonas moerusii: 単相生物Volvox aureus オオヒゲマワリ: 日本各地
雌雄異株群体 monoecisous, dioecious → 1000-3000細胞集まる ゴニジア(緑顆体) gonidium (pl. –a): an asexual, nonmotile reproductive cess, as in Volvox Volvox–細菌: 共生関係
Ex. Volvox群体中のPseudomonas fluorescens除去(無菌状態) → カバーガラスをかけ観察 HaematococcaceaePolyblepharidaceae: Polyblepharides
Class Prasinophyceae (Loxophyceae)単核Class Bryopsidophceae (Siphonophyceae)多核Subclass Hemisiphoniidae多核細胞Order Chladphorales シオグサCladophora シオグサ [f. < g. clados (枝) + phora (有する)。一列の細胞で出来た小枝を多数輪生する形状に基づいた名]C. fracta: 極めて疎らに分枝した長い糸状体。水中に浮かび枝短く1-2細胞よりなること多。細胞基部幅90-100 μm、長470-940 μm、頂で40-65 μm、250-700 μm。細胞壁肥厚し皺状に成層。道、欧州 Aegagropila linnaei Kützing (マリモ/毬藻, lake ball or moss ball), syn. Cladophora aegagropila (L.) RabenhorstOrder SphaeroplalesOrder Acrosiphoniales モツレグサAcrosiphoniaceae モツレグサUrospora penicilliformis (Roth) Areschoug シリオミドロ 潮間帯上部岩上. 北海道, 本州太平洋岸北・中部 + 海外 Subclass Cystosiphoniidae発達段階で多核。発達につれ多細胞体Order Siphonocladales
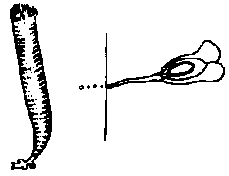
Valonia バロニア: 頭部は袋状で隔壁はない Polyphysaceae カサノリ Order Dasycladales カサノリ多核体(ケノサイト) coenocyte複相 diplont (遊走子形成なし) 輪生枝 verticillate branch + 中軸 axis
配偶子嚢 gametangium (pl. –a)
配偶体発生 gametophyte development 茎葉体 gametophore: 茎葉区別できる成長体でgametangiaを含む植物構造 Polyphysaceae カサノリ→ (細胞の)分断 merotomy Acetabularia (カサノリ)
Subclass EusiphoniidaeOrder DerbesialesOrder CodialesOrder Caulerpalesイワヅタ: クロロプラスト、ロイコプラスト、セルロース無。Xylen Caulerpaceae イワヅタCaulerpa イワヅタ [f < g caulos(茎) + erpo (地を這う)。海藻だが茎の様な部分が岩の上を長く這い廻る] コウレプシン caulerpicin (n): 有毒 Order Bryopsidales ハネモBryopsidaceae ハネモBryopsis [f < g Bryum (コケ gn) + opsis (似)。概形が細かい葉のコケに似る] Order DicholomosiphonalesClass Oedogoniophyceae サヤミドロOrder Oedogoniales サヤミドロOedogoniaceae サヤミドロ: Bulbochaete, Oedocladium, OedogoniumClass Zygonematophyceae (Conyngatophyceae) ホシミドロ鞭毛なしOrder Zygnematales ホシミドロクロロコックム目と淡水産プランクトンの主要な目Mesotaeniaceae メソテニウム |
Gonatozygaceae ゴナトヂゴン Zygnemataceae チグネマ(ホシミドロ) Zygnema Agardh ホシミドロ: Z. cruciatum Spirogyra Link アオミドロ: S. crassa フトアオミドロ Mougeotia Agardh ヒザオリ: M. scalaris ハシゴモ Desmidiaceae ツヅミモ: 高層湿原出現種多 Cosmarium ツヅミモ, Closterium ミカヅキモ Order Tetrasporales ヨツメモ藻体: 寒天状粘質に包まれた単細胞またはパルメラ状群体。細胞に偽繊毛pseudocilium持つものあり浮遊性か付着性、多くは淡水産 色素体: 杯状でピレノイドある 無性生殖: 2分裂か遊走子 有性生殖: 同型配偶子の接合 Tetrasporaceae (ヨツメモ) Collinsiella (ランソウモドキ) C. tuberculata ランソウモドキ: 単細胞緑藻だが粘質中で群体形成。その団塊はほぼ球状。潮間帯上部岩上に群生。北海道(室蘭)、表日本北部、アメリカ西岸 Palmellaceae: パルメラ状群体。偽繊毛Chlorangiaceae Order Chlorococcales クロロコックムホシミドロ目と淡水プランクトンの主要目藻体: 不動性単細胞、群体、定数群体等。浮遊性か付着性。淡水産 色素体: 杯状、星状、薄板状。ピレノイドある 遊走子・自生胞子・娘定数群体形成。同型配偶子の接合するものもある = Volvocalesと似た生殖 Chlorococcaceae Cystococcus
自生胞子嚢 autosporangium (pl. –a) Eremosphaeraceae: 稀 Chlorellaceae クロレラ Chlorella: 有性生殖欠失(核相交代、世代交代ない)
同調培養 synchronized culture Selenastraceae Dictyosphaeraceae Hydrodictiaceae アミミドロ Euastropsis Hydrodictyon アオミドロ (H. reticulatum アオミドロ) Pediastrum Sorastrum Coelastraceae Protosiphonaceae Order Ulotrichales ヒビミドロ1列細胞Suborder Ulotrichineae ヒビミドロ同形配偶 (isogamy)または異型配偶 (anisogamy)単列。無分枝。糸状体 Ulotrichaceae: Ulothrix ヒビミドロ
Schizomeridaceae Suborder Ulvineae アオサUlvaceae アオサEnteromorpha アオノリ [f < g enteron(腸) + morph (形)。葉状体作る内外2層の細胞層間に空間あり中空袋状になる]
ボウアオサ: 体筒状余り分枝ない。時に長さ数m。外洋水影響多い海岸岩上着生。邦内各地、欧州、南米等 U. compressa L. ヒラアオノリ, syn. Enteromorpha compressa (L.) Nees, green laver
潮間帯上部-中部 → 岩や藻体の上
1: 体: 全体が下部を除き大小の網目状 = アミアオサ
標本紙への付着不十分
エゾヒトエグサ: 体はかなり成長するまで嚢状をなし、後に破れる(H ≈ 10 cm)。潮間帯下部から斬深帯上部の岩上または他の海藻上に生じる。北海道、本州、大西洋
Suborder Prasiolineae カワノリPrasiolaceae カワノリPrasiola カワノリ, Schizogonium Suborder Sphaeropleineae ヨコカワミドロSphaeropleaceae: Sphaeroplea (S. annulina ヨコワミドロ, 水田産)Order Chaetophorales カエトフォラ単列糸状体 uniseriate filament。細胞に剛毛setaある原則 heterotrichous form (直立部 + 匍匐部に分化) (場合により一方のみ) 単相 Chaetophoraceae タマモ: Aphanochaete, Chaetonema, Chaetopeltis, Chaetophora タマモ, Cloniophora, Draparnaldia ツルギミドロ, Draparnaldiopsis, Endoderma, Fritschiella, Microthamnion, Protoderma, Pseudoulvella, Saprochaete (腐生藻), Stigeoclonium, Thamniochaete
Draparnaldia plumosa: 糸状体。栄養体は明瞭な主軸部と側枝部に分枝し、主軸部円柱形。細胞 φ = 30-50 μm、長50-80 μm。側枝束状、全形長紡錘形、側枝主軸明瞭。分枝対生-互生。北海道、本州 Scenedesmus sp.: 糸状。分枝あるいは分枝なし。細胞円柱状。多核 Trentepohliaceae スミレモ: 気生。生殖器官分化 Cephaleuros, Chlorotylium, Ctenocladus, Fridaea, Gomontia (G. polyrhiza 穿孔藻), Gongrosira, Leptosira, Phycopeltis, Physolium, Tretepohlia (T. aurea スミレモ) 毛 (hair) = 剛毛 (bristle hair) → 細胞壁の変形したもの Coleochaetaceae サヤゲモ: Coleochete (C. pulvinata 生殖器官分化)
Protococcaceae プロトコッカス
Order Oedogoniales サヤミドロOrder Sihponales クダモ(管状藻)(別分類: Siphonocladales, Dasycladales, Siphonalesの3つに分ける)多核体coenocyte。低温地域には少ない Order Sihpnocladales ネダシグサDerbesiaceae ツユノイト: haplodiplontDerbesia (D. lamourouxii ツユノイト): 冠毛 = 鞭毛の出方 – gameteは2本の鞭毛。異型世代交代 DasycladaceaeCodiaceae ミル (☛ Order Codiales, 見解) クロロプラスト、セルロースなし。マンナン Codium Stackhouse ミル [n < g codion(絨毛の如き皮)。又はcodeia(頭)語源。先端は頭の形をなす] |
|
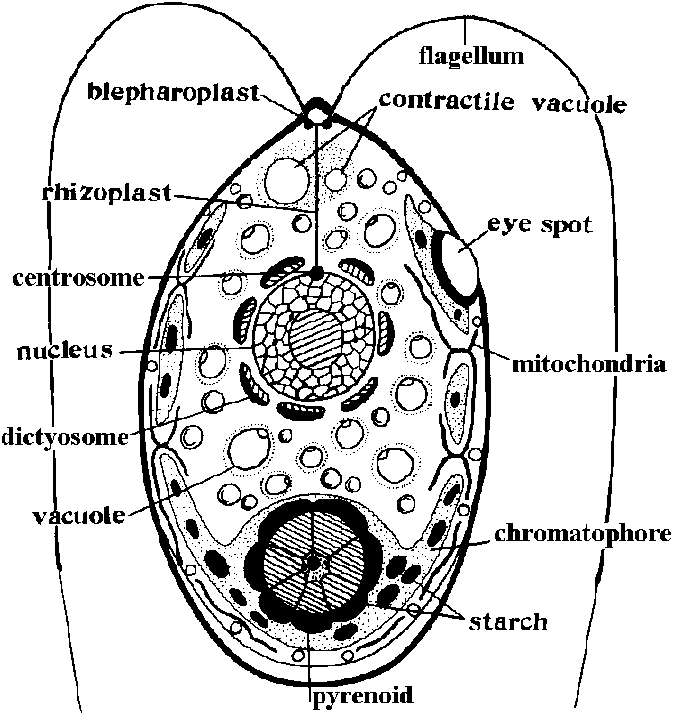
鞭毛を持つ運動性単細胞 色素体: 円盤状、薄板状、星状等でピレノイドある 葉緑体とミトコンドリアが細胞内に分散する 色素: クロロフィルa, b, カロチン、キサントフィル等 (光合成色素持たず有機栄養を営むものもある)
鞭毛基部に、特殊細胞小器官(眼点 eye spot)存在 - 特殊なカロチノイド色素を含み赤色を呈する |
Euglena gracilis Klebs (ヒメミドリムシ): 光合成鞭毛藻代表種 - 顕著な極性 細胞細長く葉状で、普通、鞭毛は不等長2本で前端部に食い込み、細胞前端部の小孔から突出 他鞭毛藻と異なり細胞は固い細胞壁で包まれない 外層: 伸縮性外皮であり、体を著しく変化させる細胞分裂: 縦分裂であり、有糸分裂が始まる頃、鞭毛とその基部構造を含む細胞の前部の細胞小器官に複製が起こり、その後縦軸に沿って分裂が起こり複製された細胞小器官は2個の娘細胞に等分される |
|
1000-1200種: 数mm - 30 m程度の個体(単細胞褐藻未確認) 同化色素: Chl a, c, β-カロチン, フコキサンチン fucoxanthin (一般にキサントフィルxanthophyllが他藻類に比べ多) 同化産物(貯蔵物質): ラミナリン laminarin or laminaran, マンニトール manitol 葉緑体: 3重チラコイドラメラ。2枚の葉緑体包膜と核膜に繋がる2枚の葉緑体小胞体を合わせ4枚の包膜 殆ど海産、汽水産、数種が淡水産。固着性。一般に寒水域に多 直立せず皮殻状のものもある。Ex. Macrocystis (米国西海岸): 30 m 褐藻分類は生活史に重点をおいて行われる → 培養実験 → 培地 (culture) uniseriate filament: 最も簡単な体制。1列細胞で普通分枝する (uniseriate, adj. 1列の、単列の) 組織偽柔組織 pseudoparenchymatous tissueか柔組織 parenchymatous tissueアルギン酸 alginic (or algin) acid: 褐藻細胞壁中層と1次膜の中にあるポリウロン酸の1種 成長 growth生殖 reproduction
無性生殖: 胞子 spore: zoospore = 不等長。側面から2本。羽型と鞭型
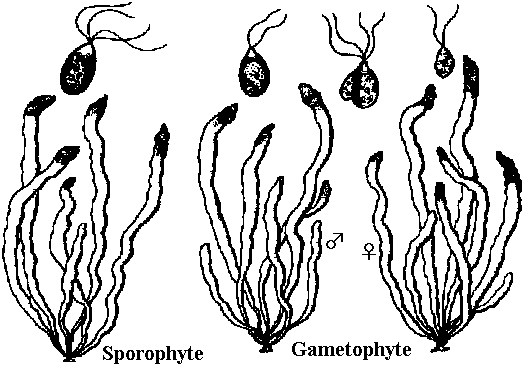
核相・世代交代haplont: なしhaplo-diplont: ヒバマタ型 isomorphic (同型): G = Sp アミジグサ型 heteromorphic (異型): G ≠ Sp
G < Sp コンブ型, G > Sp ムチモ型
→ 褐藻は緑藻に比べまとまりのよい集団 系統進化世代交代核相交代: haplodiplont: G = Sp → Sp > G → diplont
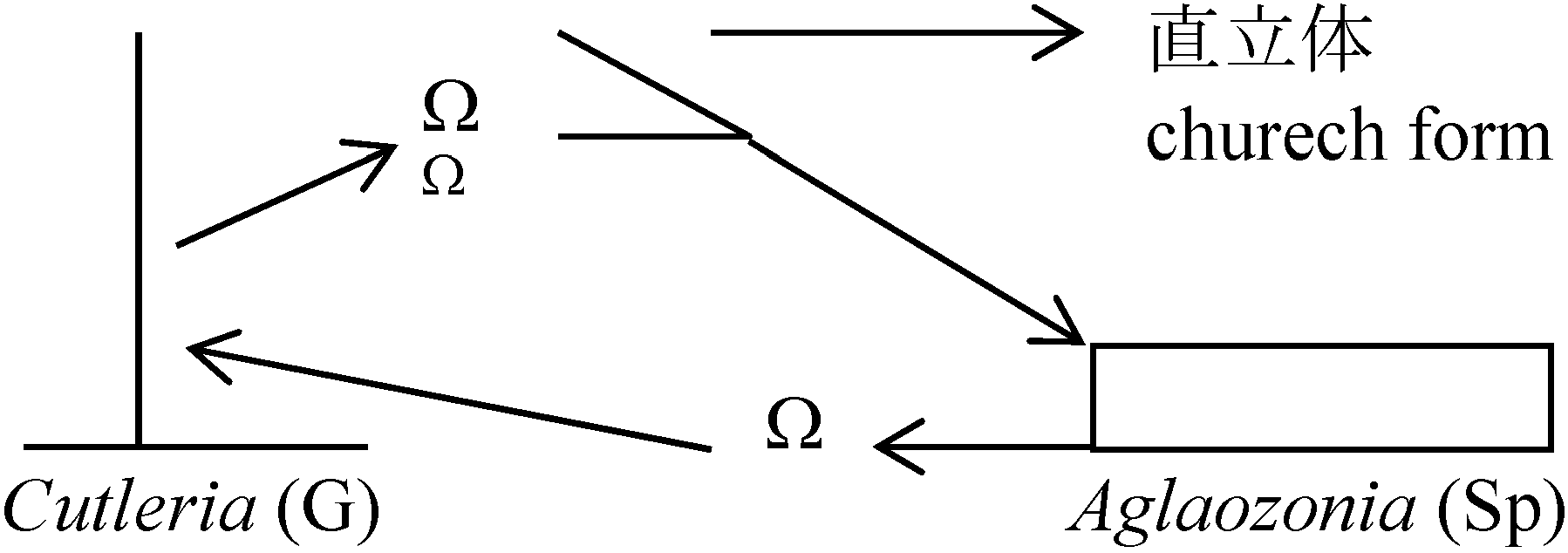
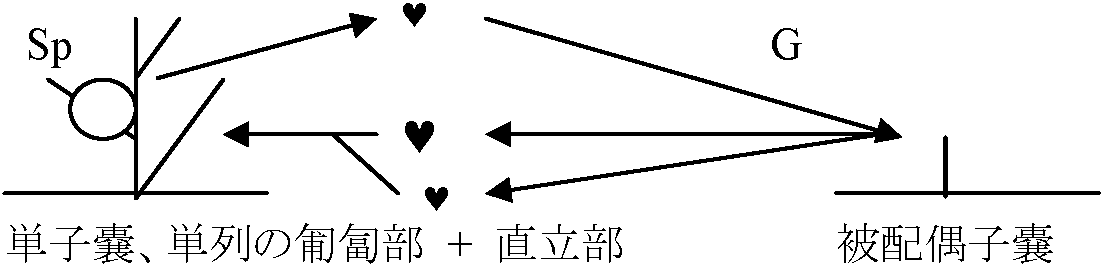
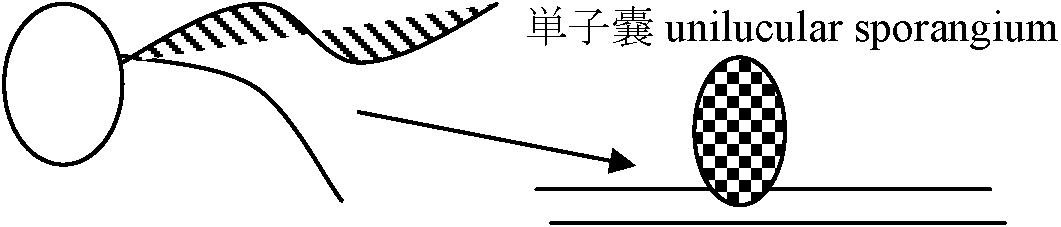
体制: 単列糸状体 → Pseudoparenchyma → Parenchyma 分類基本基準表面のつや。基部形状。先端部成長状態Class Isogenerate 同型世代鋼同型世代交代Order Ectocarpales シオミドロ単列糸状体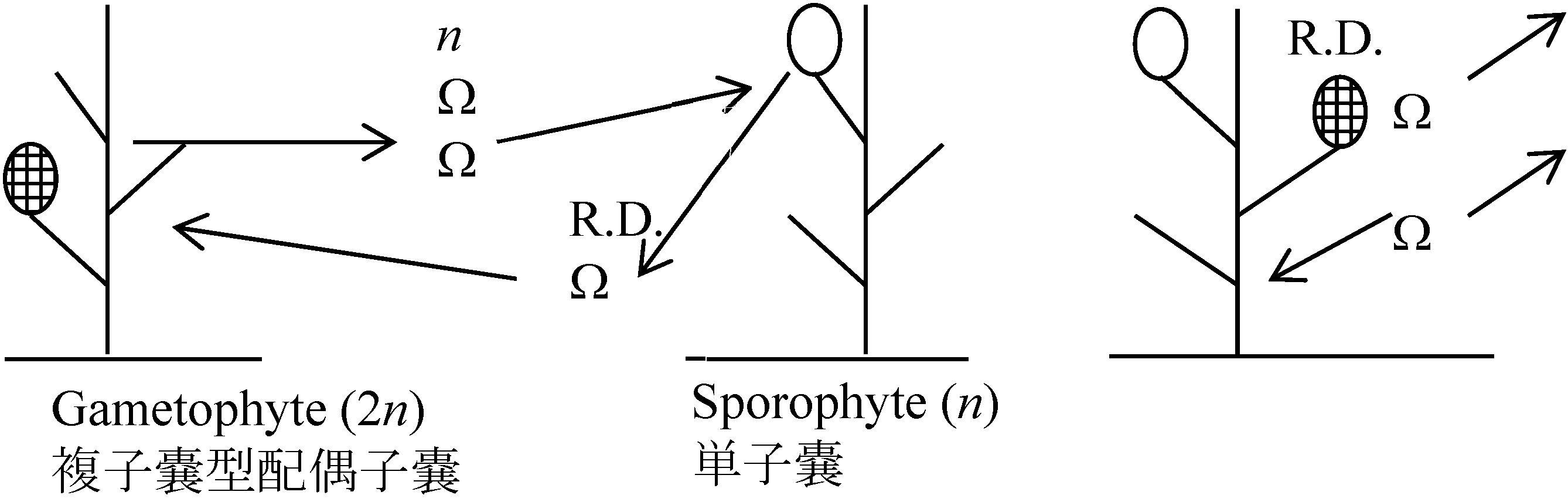 時に複子嚢型と単子嚢型の生殖器官が同一個体上に現れる 複子嚢(= sporophyte)はaccessory reproductive organで核相世代に関係なく個体増殖のためのもの Ectocarpaceae シオミドロ: Bodanella, Ectocarpus, Feldmannia, Pleurocladia, Pylaiella (P. littoralis ピラエラ: 介生的に複子嚢存在), Sorocarpus (S. uvaeformis イソブドウ) Order Ralfsiales イソガワラ(生殖 = Ectocarpales型 → Ectocarpalesに含める見解)皮殻状で岩に張り付く Ralfsiaceae イソガワラ: Analipus マツモ (= Heterochordaria [f < g+gn heteros異なった + Chordaria (gn)], = Ceratophyllum [n < g ceras角 + phyllon 葉 = 角状に割れた葉] (A. japonicus (Harvey) Wynne マツモ), Heribaudiella, Lithoderma, Ralfsia イソガワラ Order Sphacelariales クロガシラ頂端成長 apical growth – 頂端成長細胞大多列柔組織parenchyma作る 生殖・核相・世代交代 = Ectocarpales (栄養成長: 胚芽枝による) Cf. 小房 loculus, anisogamy Sphacelariaceae クロガシラ Sphacelaria californica ハネグンセンクロガシラ Stypocaulaceae カシラザキCladostephaceae Choristocarpaceae Order Cutleriales ムチモ体は柔組織を作り叉状分枝。円柱状、帯状頂毛成長 trichothallic growth, or 介生成長intercalary growth 生殖: sporophyteは単子嚢のみ。複子嚢はない。Anisogamyを行う
異型世代交代 = Sp.で核相世代に関係のない子嚢ができる個体 Cutleria [f < 人(Sidmouth) Cutler 女史名に因る。ムチモ科]Order Sporochnales (ケヤリ)胞子体は顕微鏡的で単子嚢のみ形成。配偶体 = oogamy匍匐部分がantheridiumとoogoniumに分化 → antheridiumで1細胞に1精子 雌雄同株。受精すると直立体のsporophyteになる Sporochnaceae: Carpomitra (C. cabrerae イチメガサ), Nereia, Sporochnus (S. scoparius ケヤリ) Order Desmarestiales ウルシグサ配偶体= oogamy。胞子体 = 単子嚢。中軸糸から輪生枝を出す(対生的)Desmarestiaceae ウルシグサ Desmarestia ウルシグサ [f < 人(仏学者A.G. Desmarest)]: 死ぬとH2O4放出。枝基部から糸状体皮層が中軸糸囲む。雌雄異株 Order Tiplopteridales ティロプテリス単列糸状体だが部分的に多列Haplospore(Sp): 4核の単胞子嚢monosporangia Scuphospora (G): 被子嚢 → 造精器, 1核単胞子嚢 → 造卵器 単子嚢は遊走子zoospore作る。単胞子嚢は不動胞子aplanospore作る Tilopteridacdeae: Haplosore, Tilopteris Order Dictyotales アミジグサ体2層-多層柔組織細胞か叉状分枝する偏平な直立体成長は頂端細胞分裂か縁辺細胞列marginal row of apical cells分裂のほか、介在細胞の縦横分裂も見られる。体表からは集団paranemataが点状もしくは重圏状にでる |
柔組織形成 胞子体はaplanosperangia (tetrasporangia)形成 Oogamy。Sp.で不動胞子形成はTiplopteridalesとDictyotales Apical growth: Dictyhota アミジグサ ↔ Marginal growth: Lonaria シマオウギ, Padrina ウミウチワ antheridiumとoogoniumは一個所に集まり複子嚢sorus形成 Dictyotaceae アミジグサ Dictyopteris ヤハズグサ [f < g Dictyota (gn) + pteron翼。Dictyotaに近く,然も葉中央に筋あり翼状], Dilophus ニセアミジ, Dictyota アミジグサ [f < g dictyon(網)。乾燥植物体に明瞭な網目模様がある], Distromium フタエオオギ [n < g di (2) + stroma床,層。植物体横断面細胞層2層], Homoeostrichus ヤレオオギ [m < g homoios同様 + thrix, trichos糸/毛], Pachydictyon サナダグサ, Padina Adanson [f < g padinos/pedinos扁平な生活体形] (P. arborescens ウミウチワ), Pocockiella ハイオオギ [f < gn Pocockia(属名)縮小名], Spathoglossum コモングサ [n < g spatha刀 + glossa舌。葉裂片形から], Stypopodium ジガミグサ [n < g stypos毛生えた + podus足], Zonaria シマオオギ [f < g zona重圏状線。縁に沿い筋] Dictyota dichotoma (Hudson) Lamouroux (アミジグサ): 低潮線付近に普通に分布。成実体は晩春から夏。四分胞子体多 Class 異型世代鋼 (Heterogenerate)Subclass HaplostichineaeOrder Chordariales ナガマツモ概要はEctocarpalesに似る生殖様々。単軸か多軸構造
pith, assimilatory filament に分化 ↓ 同化糸 ↓ 遊走子単子嚢のみあるいは遊走子複子嚢 その両方の3タイプ
gametophyte イシモズク Sphaerotrichia が hetero-
頂毛成長(Chordariaceae, Acrothrichaceae, Splachirididaceae) Elachistaceae ナミマクラ: Elachista, Halothrix (H. ambigua ソメワケグサ – 介生的) Leathesiaceae ネバリモ Leathesia (f<p G.R.Leathes) ネバリモ: L. difformis (L.) Areschoug ネバリモ Chordariaceae (Mesogloiaceae) ナガマツモ: Mesogloia [f < 1 mes(中間) + gloea(膠,海綿等の内胚葉と外胚葉との間の粘液)。寒天質で粘る], Eudesme [f < g eu(良い) + desme(糸束)], Sphaerotrichia [f < g sphaero(球) + trichia (糸)。中軸糸は先端に丸い細胞がある], Tinocladia [f < g tino(伸びた,拡がった) + clados (枝)。枝が拡がる] Spermatochnaceae モズク: Nemacystus [m < g nema(糸) + cystus(嚢)] Acrothricaceae ニセモズク Chordariopsidaceae Splachnidiaceae Ishigeaceae イシゲ: Ishige [f < jap イシゲ(和名)] Order Dictyosiphonales ウイキョウモ藻体: 糸状、リボン状、葉状か袋状 – いずれも多列形成的成長柔組織をなし皮層と髄層に分かれるもの多 Punctariaceae (Asperococcaceae) ハバモドキ(カヤモノリ)
→ diffuse growth Myelophycus [m < g myelos(髄) + phycos (藻)] Coilodesmaceae エゾフクロOrder Scytosiphonales カヤモノリ(Dictyoshiphonalesから独立)Scytosiphonaceae カヤモノリ: Scytosiphon カヤモノリ [m < g scytos(革) + siphon(管)。植物体が一見革質の管状呈する], Hydroclathrus [m < g+1 hydro(水) + clathratus(格子状)。水中に生じ多数の孔が空き格子状とも見える], Endarachne [< g endos(内部) + arachne(蜘蛛の糸)。植物体内部(髄層)で細長い細胞が縦に連なり,蜘蛛の糸のように絡み合う] Chnoosporaceae ムラチドリ 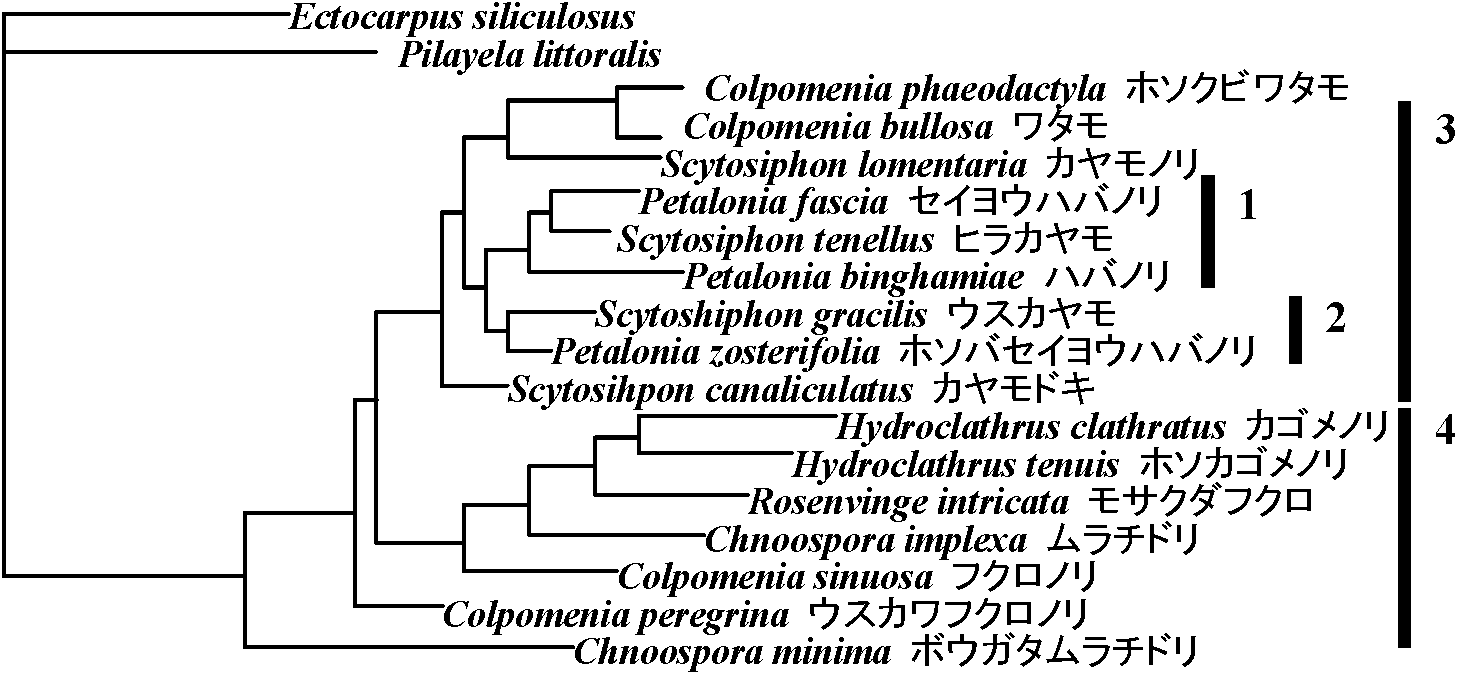 図. 分枝系統樹. 1: 胞子体はStragularia, 2: 胞子体はCompsonema, 3: 胞子体は生殖器官として端子嚢だけを形成, 4: 胞子体は端子嚢に加え副子嚢形成
図. 分枝系統樹. 1: 胞子体はStragularia, 2: 胞子体はCompsonema, 3: 胞子体は生殖器官として端子嚢だけを形成, 4: 胞子体は端子嚢に加え副子嚢形成
Subclass PolystichineaeOrder Laminariales コンブ内皮層 inner cortexから髄層 medulla に向かって糸状の突起がでる内外部の間位に粘液控導存在し粘液道 mucilage canal 基部に粘液腺 gland cell, secretary cell があり、粘質分泌する 茎葉間の meristem 部で成長 → 上部ほど古く、秋には葉 blade 上方枯れる 冬に成長帯が分裂を開始し、古いbladeを持ち上げるように再生してくる 単子嚢: 胞子葉 sporophyll という特別な葉にのみできる 雌 gametophyto 上に sporophyte できる。雌雄配偶体の大きさが全く異なる Laminariaceae コンブ 卵が移動しないのが特徴  ● ホソメコンブ
● マコンブ
● リシリコンブ
● オニコンブ
● ナガコンブ
● ミツイシコンブ
● ホソメコンブ
● マコンブ
● リシリコンブ
● オニコンブ
● ナガコンブ
● ミツイシコンブArthrothamnus ネコアシコンブ (m < g arthron (関節) + thamnos (低木,枝)。葉が広い枝状に見え,旧葉は関節で落ち去る) Costaria スジメ (f < l costa(中脈)。葉面に太線条目立つ) Laminaria コンブ (f < l lamina(葉)。植物体大きな葉状) ホソメコンブ L. religiosa Miyabe: 実験室培養条件: 14/10hr (1500 lux), PES培地 Undaria (f < 1 unda 波,流水,潮)Ecklonia (f < p アフリカ植物研究者C. F. Ecklon, 19C) Eisenia (f < p Eisen博士) Chordaceae ツルモ Chorda Stackhouse ツルモ (n > l chorda(紐)。体形状から) (C. filum ツルモ) Alariaceae チガイソ Alaria チガイソ (f > l ala 翼。体は中肋の両側に広く翼状に拡がる) Lessoniaceae レッソニア Class Cyclosporae (円胞子鋼)全て複相胞子世代diplontで、単相配偶世代は独立した藻体では存在しないOrder Fucales ヒバマタ体中-大型(4-5 mに達する)頂端成長 apical growth。無性生殖ない oogamy antheridium 64, oogonium 8核 (8, 4, 2, 1と卵の数は不定), 卵・精子だけn 生殖器巣 conceptacle Ascoseriaceae Durvilleaceae Notheiacaee Fucaceae Anderson, 1763 ヒバマタ Fucus L. ヒバマタ (m < g Grで海藻を指す言葉phycos由来): 受精卵は極性研究に古くから利用(光照射方向、電流、水素イオン濃度勾配、遠心等) → 形態形成・ホルモン (F. evanescens ヒバマタ) Pelvetia Deéne (f < p 仏19C自然愛好家Pelvet) (P. wrightii エゾイシゲ), Phyllospora, Scytothalia (Ascophyllum nodosum (L.) Le Jol), [Cystophyllum n < g cystis 気胞 + phyllon 葉。葉先に気胞あり] Himanthaliaceae Cystoseiraceae Sargassaceae ホンダワラ Coccophora, Hizikia Okamura [f < jap ヒジキ(日)] (H. fusiforme ホンダワラ), Sargassum Agardh [n < sp 海藻(sargago, 西語)由来。大西洋”藻の海”の主体] (S. fulvellum), Turvinaria |
棹前昆布
種類 棹前早煮昆布3等 (300g) 棹前早煮尺頭1等 (100g) 棹前昆布は、7月から採取されるなが昆布の一部を充分身入りしない若い状態で採取される昆布であり、2年目の昆布が中心である。 採取される期間は6月中旬-6月下旬までの短期間であり、近年ではその短期間の中でも5日間程度で1日2時間程度の操業しかせず、生産量も限られている。採取された時の幅は6-10cm程である。 代表的なシーベジタブル(海の野菜)と言われるほど葉質は柔らかくなが昆布地帯だけで獲れる特別な昆布である。又、生育密度が非常に高いのも特徴であり、なが昆布の生育を促す為には好条件な漁業と言えるだろう。 漁具は「鈎」と言われる木製の棹であり、棹前昆布の名前の由来は、なが昆布の棹を立てる前に採取する昆布と言う意味であると言われている。 採取された棹前昆布は手間暇かけて乾燥させ、根(先端部分50cmか105cmにカット)・葉(先端の次より下を105cmにカット)に切り分けられ葉幅・色目・長さ・量目により等級分けされ、全国に出荷される。 豆知識: 通常の昆布は根をダシとして主に利用するが棹前昆布は利用しない為、50 cm以上に切られる。 ■■■ 分 布 ■■■ 釧路港以東、昆布森、厚岸、浜中、散布、花咲を経て歯舞に至る太平洋沿岸、貝殻島、歯舞諸島、クナシリ島、エトロフ島 ■■■浜の母さん情報■■■ 棹前昆布を使う定番家庭料理は、結び昆布・おでん・煮物・昆布巻・サラダ。調理が手軽な昆布で、用途様々に使用でき、お勧めな昆布。結び昆布に最適で、圧力鍋で15分程度(普通鍋で40分程度)で煮える。大根等の山の幸と一緒に炊く事でより早く煮える。 ● 結び昆布の作り方 □□□材料□□□ 棹前昆布…100 g、酒…大匙5杯、砂糖…大匙2杯、醤油…大匙4杯、味醂…大匙3杯、酢…少々
1. するめは薄皮を剥き、3-4cmの長さに揃えハサミで細切りにする 2. 酒を温めてするめにかけて軟らかくする 3. 昆布を濡布巾で拭き、するめと同様細切りにし、酢をかけておく 4. 人参は3-4cmに千切りし、赤唐辛子は種を抜き小口切りにする 5. 数の子は、一晩水につけて塩抜きしてから、小さくほぐす 6. 鍋に、★を合わせて入れ、火にかけてひと煮立ちさせる 7. 漬け容器に2-5を入れ、6を注いで混ぜ合わせる 8. 皿などで軽く重石をかけ、ときどき上下を返して混ぜる 9. 1日くらい漬け込み出来上がり |
豆知識: 醤油ご飯 - 細かく刻んで一緒に炊くだけで風味抜群。湯豆腐 - 豆腐と同じ位に切って、サッと煮れば一緒に食べられる。シチュー - 小さく切って、出来上がる寸前に入れるだけで美味しい。
なが昆布
種類 長昆布1等 (300 g) 長昆布1等元 (300 g) なが昆布は、文字通りに非常に長い昆布で、長さは平均で7-8m、長いものは20mにも達し、世界の昆布中最も長い昆布である。 採取時期は7月初旬-9月下旬までで、採取日数は近年20-25日程度である。この昆布の分布範囲は決して広いとは言えないが、成育密度が高く、資源量は他の昆布地帯に比べ格段に安定していると言える。なが昆布の寿命は発芽時期によって変わり、夏場に出た芽は3年生、冬場に出た芽は2年生葉体になるが、高密度や日光不足で成長が抑えられると、1年採取時期が遅れる事がある。形状は三石昆布に良く似て細長く、中帯部も狭い溝状になっている。 漁具は「鈎」と言う木製の棹と「ネジリ」と言うコンポーズパイプであり、この2つの使い分けは、通常資源量の多いお盆前までは「鈎」で昆布をすくい上げ、お盆以降は短い昆布が主となる為、すくい上げる事が出来ず海中で昆布を挟んでねじる「ネジリ」を使用する。採取された昆布を手間暇かけて乾燥させ、根(先端部分15-20 cm)・元(先端の次、一番切り105 cm)・葉(二番切り以降105 cm)に切り分け、葉幅・色目・長さ・量芽により等級分けされ全国出荷される。 ■■■ 分 布 ■■■ 釧路港以東、昆布森、厚岸、浜中、散布、花咲を経て歯舞に至る太平洋沿岸、貝殻島、歯舞諸島、クナシリ島、エトロフ島 ■■■浜の母さん情報■■■ 身入良く、基本的にダシ(おでんに最適)や昆布巻・佃煮等に利用する。結び昆布等で硬いものを好む方は、長昆布で調理すると歯応え残り満足できると思う。おでんのダシをとった後に召上がるのも美味しい。小さく切って醤油さしに入れておけば醤油にダシが溶け出し円やかになる。 ● 昆布しゅうまいの作り方 □□□材料□□□ はんぺん…1枚、なが昆布…1枚、たけのこ…1/4個、しいたけ…1枚、シュウマイの皮…5枚、塩・胡椒…少々、人参…1/4個、長ネギ…1/4個
1. 昆布は水に戻し、4等分する 2. 1を繊維に直角に屏風折にし、ベーコンを巻いて爪楊枝で止めておく 3. フライパンで2を焼く 4. 鍋に水・コンソメ・塩・黒胡椒を入れ火にかけ沸騰したら3を入れ煮る 5. 昆布が軟らかくなったら一度取り出し、とき卵流し込み、火止めベーコンを巻き昆布を戻す 6. スープ皿に5を入れ、パセリを散らし完成 |
|
種類 あつば昆布1等検 (200 g) あつば根昆布 (100 g) あつば昆布■■■ 特徴 ■■■別名ガッカラコンブ。あつば昆布は、文字通り葉質硬く生の時は基部の中帯部を二つ折にすると音を立て割れるほど脆い昆布で、「ガッカラ」とは乾燥葉が硬い事に由来する。道東では、なが昆布が有名な為に他地方では馴染みが薄いが、価格では近年なが昆布以上の評価を得る。 長さ2-5mで幅8-20cm程度と厚みがある。基部円形、中帯部広く全幅の2/3を占める。分布域はなが昆布と全く同じだが、暗礁の陰や海底の凹地など波の当たらない深みを成育場所とし、波の荒い場所を好むなが昆布と生活領域を分け合う。全長が短い為採取漁具は「ネジリ」と言う海中で昆布をねじるものを主に使用する。 採取された昆布を手間暇かけ乾燥させ、根(先端部5-8cm程度)・葉(先端の次1番切り105cm)に切り分け葉幅・色目・長さ・量目により等級分けされ全国出荷される。根昆布は昆布水用として根強い人気商品である。 ■■■ 分 布 ■■■ 釧路、昆布森、厚岸、散布、浜中から納沙布岬に至る各地、貝殻島、色丹島、クナシリ島 ■■■浜の母さん情報■■■ 身入り良く幅広い昆布で、一番利用が高いのは昆布巻。さんまや鮭と一緒に昆布巻にすると美味。ダシや佃煮等にも利用出来る。高血圧や便秘の方は根昆布を昆布水にし毎日利用する方が多く、あつば昆布でなければダメという声も。油で揚げグラニュ糖を付けお八つにもなる! ● 昆布しゅうまいの作り方 □□□材料□□□ あつば昆布…70g、秋鮭…400g、酒…2/3カップ、砂糖…大匙5杯、醤油…1/2カップ、酢…1/4カップ 1. 昆布は8カップに10分程度浸しておく 2. 戻した昆布を水から上げ、鍋幅より短めに切る 3. 秋鮭の皮骨を除き、3cmの棒状に切り昆布で巻く 4. クッキングペーパーを細く切ったもので所々結ぶ 5. 鍋に昆布の戻し汁・昆布巻き・酢・酒を入れ30分位煮る 6. 砂糖と醤油を入れ、とろ火で1時間ほど昆布が軟らかくなるまで煮込む 7. クッキングペーパーをとり、食べやすい大きさに切る ● あつば昆布の佃煮の作り方 □□□材料□□□ あつば昆布…100g、醤油…1/2カップ、砂糖…1カップ、酒…少々1. 昆布を水で戻し千切りにする 2. 昆布と他の材料を、昆布が隠れる程度鍋に水を張り入れる 3. 弱火で煮汁がなくなるまでコトコト煮て出来上がり ● 昆布水の作り方1. コップ6分目位水を入れる 2. 根昆布を2枚分程度入れる 3. 24時間ダシを出し完成 豆知識: 毎日続ける事で高血圧・便秘・美容に効果がある。おに昆布(鬼昆布)種類 厚岸鬼昆布1等検(200 g) おにおぼろ(30·50g) おにとろろ(80·160g) ■■■ 特徴 ■■■ |
鬼昆布は道東部太平洋沿岸各地や根室海峡に分布し、湾内・入江・島陰・港内など波浪が直接当たらない静穏な場所で生育するため葉体は大型になる。形状は、典型的に茎が太く、短く葉幅広く基部円く張出し中帯部広く、基部付近厚さは5 cm以上になる程厚いが、縁辺は比較的薄く大きく波打ち重なり合う昆布であり羅臼昆布と同種類の昆布である。通常は長さ1.5-3mで幅20-30cmで中帯部は厚く5mm以上にもなる。 漁業道具は海中で昆布を挟みねじる「ネジリ」を使う。採取昆布は乾燥させるが、厚みのため、相当の手間暇がかかる。又、製品にする際に丸まる昆布を伸し平らにし、色目で赤・黒に分け、葉幅・量目・長さにより等級分けされる。 香り良い最高級だし昆布とし主に利用され、黄色味帯びたコクのあるだしは高い評価を得ており、昆布茶等の加工食品にも使用される希少な昆布である。 ■■■ 分 布 ■■■ 厚岸・霧多布・根室半島周辺・羅臼沿岸・クナシリ島・エトロフ島・歯舞諸島 種類 ねこ足昆布とろろ(80·160g) ねこ足昆布おぼり(40·80g) ねこ足昆布■■■ 特徴 ■■■名の由来は昆布の茎とそれから出る根の様子がねこ足の様に見える事から。この昆布は外洋に面した岩礁域で水深5-7 m程の海域に群生し、岸部や船上から生育状況確認は難しい。しかし、厚岸大黒島では干潮付近の岩礁に着生するのが見られ、珍しい海域と言える。 特徴は、長さ2-3mで幅5-7cm程度、色暗褐色で乾燥すると黒色になり、葉質は硬い革質である。変わった特徴としては、1年目は他の昆布と変わらないが、2年目の再生時は葉の基部の両側の耳全体からそれぞれ新しい茎を持つ新葉が出来、やがて中央の旧葉は流れ去り、葉体を見ると、まるで古い茎が2股状に分かれ2枚の葉を持った昆布に見える。寿命長く、4-5年までは成育が確認される。マンニットを多量に含み、とろろ分と甘味に優れ、葉の状態で流通する事は殆どなく加工用として評価の高い昆布である。 ■■■ 分 布 ■■■ 昆布森・厚岸・散布・浜中から納沙布岬に至る太平洋沿岸、歯舞諸島、色丹島、クナシリ島、エトロフ島 種類 とろろ刻み昆布 (50g) とろろ足昆布■■■ 特徴 ■■■とろろ昆布は僅か2種類からなる小さな属で、北海道を中心に極東地域にのみ分布し、中でも狭い海域にしか分布が見られず希少な昆布である。特徴は狭く平坦な中央部と、その両縁の全面に形成された雲紋上の凹凸である。この模様は細かく一つ一つが整い水平に並んでいる。但し、乾燥すると凹凸模様は不明瞭になる。 長さ1-2mで、幅7-15cmに達する。粘質が極めて多い特徴を持ち、葉の状態の流通ではなく加工品の原料として扱われる。 ■■■ 分 布 ■■■ 三ツ浦・又飯時・厚岸・散布・浜中から納沙布岬に至る沿岸、歯舞諸島・クナシリ島・エトロフ島・根室港外周辺 |
|
600 gn., 6000 spp. 大部分海産で淡水産少 核、葉緑体、ミトコンドリア等の分化器官形成 同化色素 生殖・生活史極めて複雑。生殖細胞鞭毛持たず、運動段階mobile phaseない造果器 carpogonium の果胞子 carpospore 形成までが重要な分類基準 無性生殖
単胞子 monospore
卵生殖 oogamy: 卵egg, oogonium + 不動精子spermatium, antheridium or spermatangium
1重チラコイドラメラ セルロースかβ-1:3キシランとβ-1:4マンナン 系統 (再編必要)Phylum CyanidiophytaPhylum RhodophytaClass Protoflorideae (Protoflorideophyceae) 原始紅藻鋼, orClass Bangioideae (Bangiophyceae) ウシケノリ鋼= 造果器 carpogonium 中に果胞子をつくる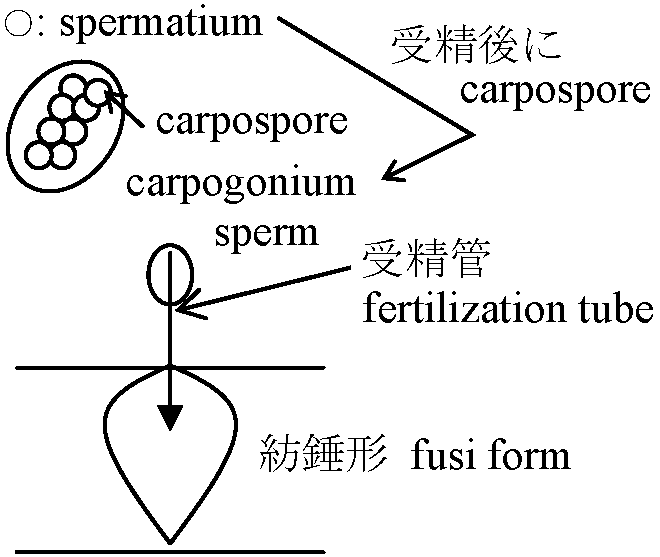
夏のり説: 直立し発達するのが一般的 夏期生活形態(説)
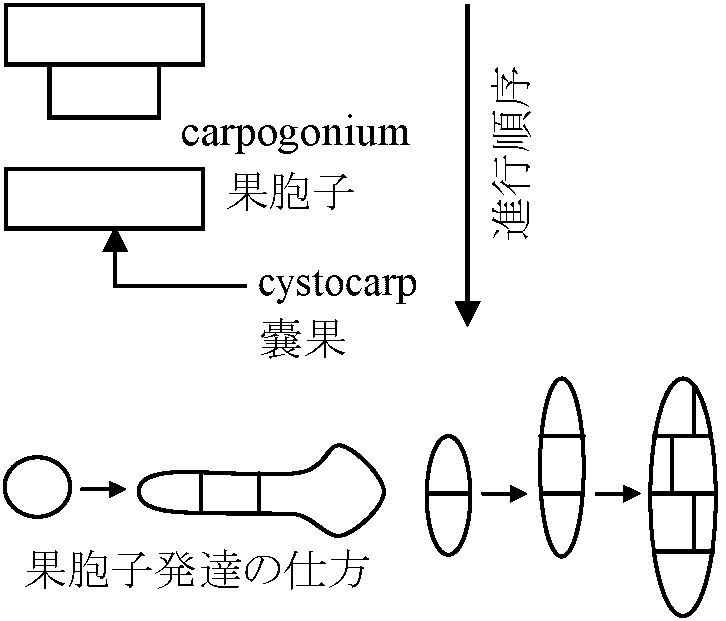
無性生殖: 単細胞 – 2分裂、多細胞 – 単胞子 (有性生殖はウシケノリ型では一部分が行うにすぎない) Order Porphyridiales チリノモ Porphyridiaceae: Chroothece, Porphyridium Order Goniotrichales ベニミドロ Order Bangiales ウシケノリ Erythropeltidaceae: Colathrocladia, Erythrocladia, Erythropeltis, Kyliniella, Membranella, Porphyropsis, Smithora Bangiaceae ウシケノリ Bangia ウシケノリ (f. < 人(植物学者Hofman-Bang), Erythrotrichia, Porphyra アマノリ (f. < g. porphyr(紫色)に由来。植物体の色から) (P. tenera アサクサノリ, 果胞子石灰中に潜る, P. yezoensis スサビノリ) 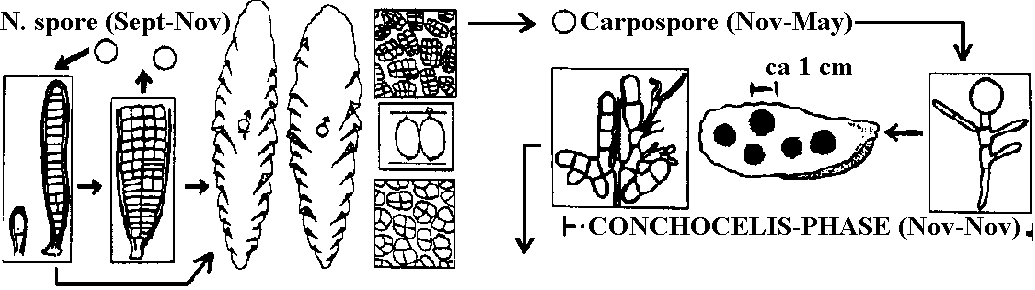 _↑----------------------------------------- ○ Meiospore (Sept-Nov) _--------------- Sept-May -------------- Fig. Porphyra: 雌雄同株と雄株の2型個体がある = androdioecious Order Compsopogonales オオイシソウ Compsopogonaceae: Compsopogon Order Rhodochaetales ロドケーテ Rhodochaetaceae: Rhodochaete Class Floridephyceae 真正紅藻鋼造果器: 造胞糸(造果糸) gonimoblast (filament)という単列糸状体を作り、これに果胞子をつくる= 真正紅藻に見られる雌性配偶子嚢 造胞糸: 最先端にだけ果胞子嚢作り、そこに各々1果胞子形成多くは成長に階層性存在: 1軸成長 > 2軸成長 > … 形態・成長 藻体: 単細胞、1細胞列、大部分は多細胞の糸状体、細状体あるいは多細胞葉状体 外見 = 葉状部 lamina + 茎状部 stipe + 付着部 holdfast 内部 = 皮層 cortical layer + 髄層 medullary layer (medulla) 成長: 単軸型か多軸型 Order Nemalionales (Nemaliales) ウミゾウメン 造果糸 gonimoblast: 受精した造果器、造果器の老細胞、あるいは器下組織 hypogynous cellから出される 嚢果 cystocarp: 紅藻類の配偶体に形成される一種の生殖器官。造果枝から発達した造果糸や果胞子の集塊。特別な助細胞 auxiliary cell ない 体制: 単列細胞による糸状体形成、単軸-多軸構造がある。原形質連絡糸に第1次連絡糸と第2次連絡糸核相交代・世代交代: 複相 haplodiplont - 配偶体と胞子体とは同型か異型 (a) 単軸型 uniaxial form Acrochaetiaceae (Chantransiaceae) Acrochaetium, Rhodochorton 単列 Batrachospermaceae カワモズク Batrachospermum カワモズク: 四分胞子体未確認。種毎に受精毛形・サイズ異 (壺形 B. ectocarpum, 徳利形 B. moniliforme, 円柱形 B. testale, B. virgatum, B. coerulescens, 瘤付形 B. graibussoniense, 倒円錐形 B. vagum, 棍棒形 B. gallaei) Lemaneaceae Lemana 果胞子発芽体が途中で減数分裂を体細胞上で行う (Batrachospermaceae同様) Naccariaceae Naccaria Bonnemaisoniaceae Asparagopsis (Falkenbergia), Bonnemaisonia (Hymenoclonium, Trailliella), Delisea, Ptilonia 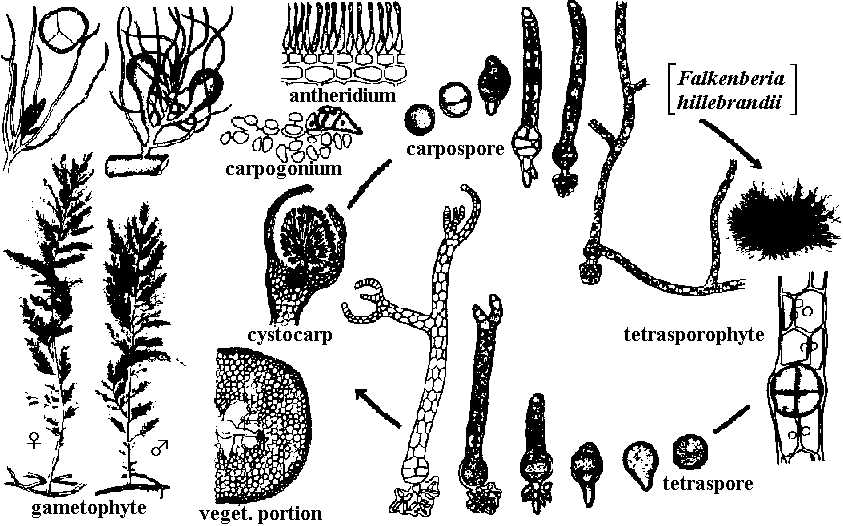 Fig. Asparagopsis taxiformis (b) 多軸型 multiaxial form Thoreaceae (Nemalionopsis), Thorea Helminthcladiaceae ベニモズク Cumaqloia, Dermonema, Helminthocladia (f < g helmins 蠕虫 + clados枝 = 虫様な枝), Helminthora, Liagora, Liagoropsis, Nemalion (n < g nema 糸。糸状植物体から), Nemalinopsis, Trichogloea Chaetangiaceae ガラガラ Actinotrichia (n < g aktis 放射線 + trix 毛。体表全体に亘う短硬毛が短い間隔で放射状輪生), Chaetangium, Galaxaura (f < g gala 乳 + auros 黄金。体に沈殿する石灰を白金と洒落た?), Gloiophloea (f < g gloios 粘質 + phloios 皮層), Scinaia (f < 人 D. Scina) 真正紅藻類生活環 (life cycle)多くは3形態相持つ
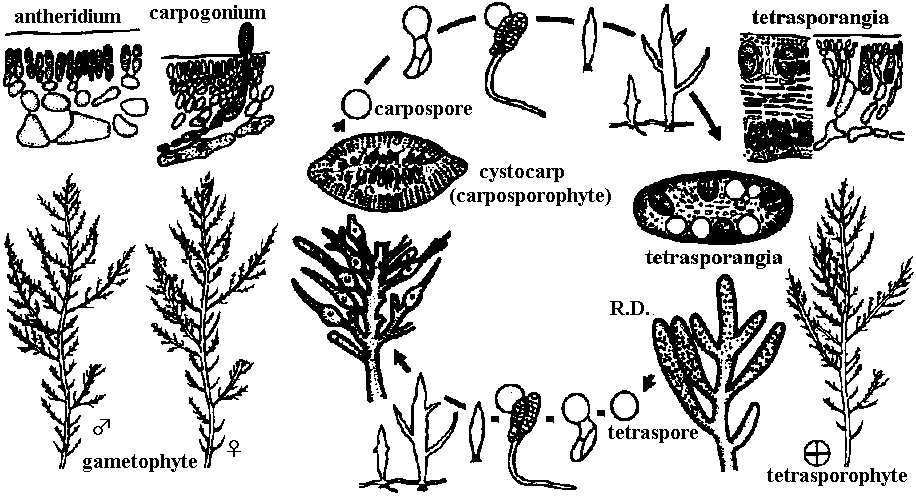 Fig. Gelidium amansii: 枝先端に嚢果形成 |
受精後、造果子は造果器から直接出る。特別な助細胞ない。一方、側枝基部から栄養細胞出され、これらの造胞子と連絡し栄養補給を行い、造胞子上に果胞子形成 体: 全て単軸構造 四分胞子嚢 teterasporophyte: 全て十字状分裂 Gelidiaceae テングサ Acanthopeltis [f < g akantha(刺) + peltis(楯)。楯状の小枝に刺を持つ], Gelidiella, Gelidium テングサ [n < 1 gelu(粘質)。粘性ゼラチン状物質(寒天)を採るため], Pterocladia [f < g pteron(翼) + clados(枝)。羽状に付く枝の状態から], Suhria [f < p 南アフリカ海藻を調べた J.N.Suhr記念], Yatabella Gelidium amansii マクサ: トコロテン・寒天材料
トコロテン: 鍋で煮て常温で固めたもの 造胞子形成し、その際、助細胞 auxiliary cell 存在し、助細胞には特別に1本の枝が出てそれが当たる。時に造果枝 carpogonial branch、支持細胞 supporting cell が助細胞になる。受精した造果器が助細胞に連絡する際、直接の場合と造果枝の1つの細胞と融合し連絡する場合がある 藻体: 単軸型/多軸型。果胞子形成はカクレイト型またはその変形 体構造: 単軸/多軸構造 四分胞子: 十字状(Ex. ヘラリュウモン、ニセカレキグサ)と平行状(アカバ) Dumontiaceae ヘラリュウモン: Counstantlnea, Cryptosiphonia, Dudresnaya, Dumontia ヘラリュウモン (D. simplex ヘラリュウモン: 体葉状、単状。10-30 cm h, > 3 cm w。潮帯間岩上に生ずる。表日本北部、北海道北岸), Farlowia, Hyalosiphonia, Neodilsea, Pikea Rhizophyllidaceae ナミノハナ: Chondrococcus [m < g chondros軟骨 + coccos 小粒], Contarinia, Rhizophyllis, Rhodopeltis Squamariaceae: Cruoriopsis, (Hildenbrandtia), Peyssonnelia (Rhododermis) Hildenbrandtiaceae: Hildenbrandtia Corallinaceae サンゴモ: 体壁に石灰質を厚く沈着する a) Amphiroideae カニノテ: 粘質栓持つ。細胞融合なく2次的原形質連絡欠く。膝節生じない Clathromorphum, Dermatholithon, Fosliella, Goniolithon, Hydrolithon, Lithophyllum, Lithoporella, Lithothamnion, Mastophora, Melobesia b) Corallinoideae サンゴモ: 粘質栓欠く。細胞融合あり2次的原形質連絡ない。膝節生じるAmphiroa [f < g amphi周囲 + roa漏れる], Calliarthron, Cheilosporum, Corallina, Jania, Joculatoa, Pachyarthron, Yamadaia Grateloupiaceae ムカデノリ: 近縁科区別点 = 造果枝と助細胞は、それぞれ別々のアンピュラ (ampulla, ローマ時代の壷)というフラスコ状細胞糸上に形成Aeodes [f < g 体葉状に広がり柔かく羊皮様], Carpopeltis [n < g carpos(果実) + pelte (楯)], Cryptonemla, Cyrtymenla, Grateloupia [f < p, 海藻学者J.P. Grateloup], Halymenia [f < g hals(海) + hymen(膜)。体膜質で海産], Pachymenia, Pachymeniopsis, Phyllmenia, Polyopes, Prionitia Gloiosiphoniaceae: Gloiosiphonia, Schinunermania (Baylesia)Endocladiaceae: Endocladia Tichocarpaceae: Tichocarpus Callymeniaceae ツカサノリ Callophyllis [m < g callos(美しい) + phyllon (葉)。葉状体紅色で美しい], Callymenia, Cirrulicarpus, Erythrophyllum, Microcoelia (Cruoriaceae): Cruoria, Petrocelis Order Gigartinales スギノリ 助細胞枝carpogonial branchなく、助細胞auxiliary cellは体構成する普通な細胞枝叢の中肝細胞のどれかが特別に栄養を貯え大きくなる。カルポゴンcarpogoneは極短いカルポゴン枝頂端にあり接合枝は助細胞へ連絡糸を出し癒合し、その助細胞から造胞糸を体内か体外へ向かい出し嚢果形成。四分胞子嚢は皮層中に存在するか特別な枝に群生、即ち、藻体皮層部糸状細胞列の1つが助細胞に変わり、それと造果器から伸びた連絡糸が連絡。後に助細胞は分裂し果胞子形成 Calosiphoniaceae: Bertholdia, Calosiphonia Nemastomaceae: Nemastoma, Platoma, Schizymenia Sebdeniaceae: Sebdenia Gracilariaceae オゴノリ Ceratodictyon (Marchesettia), Corallopsis, Gelidiopsis, Gracilaria [f < 1 gracilis(細い)。細く糸状の外形に基づく], Tylotus Gracilariopsis [f < 人 (Gracilaria(属名) + opsis (似))。即ちGracilariaに似たもの] Plocamiaceae: Plocamiocolax, Plocamium Sphaerococcaceae: Caulacanthus, Phacerocarpus, Sphaerococcus Stictosporaceae: Stictosporum Sarcodiaceae: Chondrymenia, Sarcodia, Trematocarpus Furcellariaceae: Furcellaria, Hararachnion, Neurocaulon Solieriaceae トサカノリ: Agardhiella, Euchenma [n < g eu良い+cheuma溶解するもの], Meristotheca トサカノリ [f < g meristos分裂 + thece鞘,小室], Solieria エゾナメシ, Turnerella Rissoellaceae: Rissoella Rhabdoniaceae: Catenella, Rhabdonia Rhodophyllidaceae: Acanthococcus, Cystoclonium, Rhodophyllis Hypneaceae イバラノリ: Hypnea [f < g Hypnumに似る点から付いた名。hypao(眠り)] Mychodeaceae: Mycodea, Ectoclinium Dicranemaceae: Dicranema Acrotylaceae: Acrotylus, Hennedya Phyllophoraceae オキツノリ Ahnfeltia [f < 人(Ahnfelt)], Ceratocolax, Gymnogongrus [m < g gymnos(裸) + gongros (節)], Phyllophora, Stenograrnme Gigartinaceae スギノリ: Chondrus [m < g chondros軟骨。体が軟骨質], Gigartina [f < g gigarton 葡萄種子。嚢果形状がブドウ種子に類する種がある], Iridaea [f < g iris 虹。体がiridescence(蛍光)を発するから?], Iridophycus, Rhodoglossum Gloiopeltis [f < g gloios粘質 + 属名Carpopeltisの一部分。外形 Carpopeltisに似るが, 粘質を出すことに基づく名] Order Rhodymeniales ダルス 造胞子: 助細胞から形成。助細胞は造果糸支持細胞からでた細胞がなり、これは造果器が受精前からできる。体構造は全て多軸構造。四分胞子は十字状(Ⓧ)か三角状に分裂 Rhodyrneniaceae ダルス Botryocladia, Chrysyrnenia, Coelarthrum [n < g coelos(中空) + arthron (関節)。円柱形で中空の植物体が所々で著しく括れ関節状], Cryptarachne, Erythrocolon, Fauchea, Gloioderma, Halosaccion, Rhodymenia, Weberella
Rhodymenia ダルス Order Ceramlales イギス 造胞子: 助細胞から形成。この細胞は造果器が受精した後、造果枝から出される 体構造: 単列糸状体か単軸構造(多軸構造はなし) Ceramlaceae 単列糸状体/単軸構造 Acrothamnion, Antithamnion, Callithamnion, Campylaephora [f < g campylos湾曲 + phora持つ。枝先鈎状に曲がる], Carpoblepharis, Centroceras, Ceramium イギス [n < g ceramion 壷/ceras角。枝角状に尖るから?], Crouania, Dasyphilla, Delesseriopsis, Euptilota, Griffithsia, Microcladia, Monospora, Neomonospora, Platythamnion, Pleonosporium, Plumaria, Plumariella, Ptilota クシベニヒバ, Rhodocallis, Seirospora, Spermothamnion, Spyridia, (Trailliella), Wrangelia Delesseriaceae コノハノリ Acrosorilum, Apoglossum, Branchioglossum, Caloglossa, Congregatocarpus, Cryptopleura, Delesseria, Erythroglossum, Hemineura, Holmesia, Hymenena, Hypoglossum, Hypophyllum, Laingia, Martensia, Membranoptera, Myriogramme, Nienburgia, Nithophyllum, Phycodrys, Polyneura, Pseudophycodrys, Sarcomenia, Schizoseris, Taenioma, Vanvoorstia [f < 人 (Joh.van Voorst)], Yamadaphycus Rhodomelaceae フジマツモ Acanthophora, Acrocystls, Arnansia, Aneura, Bostrychia, Chondria [f < g chondros軟骨。体が軟骨質], Digenea [f < g dis 2つ + genea属] (D. simplex マクリ → 駆虫剤), Enantiocladia, Enelittosiphonia, Euzoniella, Herpopteros, Herposiphonia, Isoptera, Laurencia, Leveillea, Neuryrnenla, Odonthalis, Polysiphonia, Pterosiphonia, Rhodomela, Roschera, Symphyocladia [f < g symphyes 結合する+ clados 枝], Vidalia, Wrightiella Polysiphonia akkesiensis Segi DasyaceaeBenzaitenia, Dasya, Dasyopsis, Heterosiphonia, Thuretia
普通は単軸的 monopodial branching であるが Dasyaceae は連基的 sympodial branching である 磯焼現象無石灰藻(サビ亜科, モカサ、キタイシモ等)、有節石灰藻(ピリヒバ、イソキリ等)増加し海底に他藻類が固着できない状態 → 悪環境のため他海藻生息できず、範適応性石灰藻が優占する(説) (否定見解もある) |
|
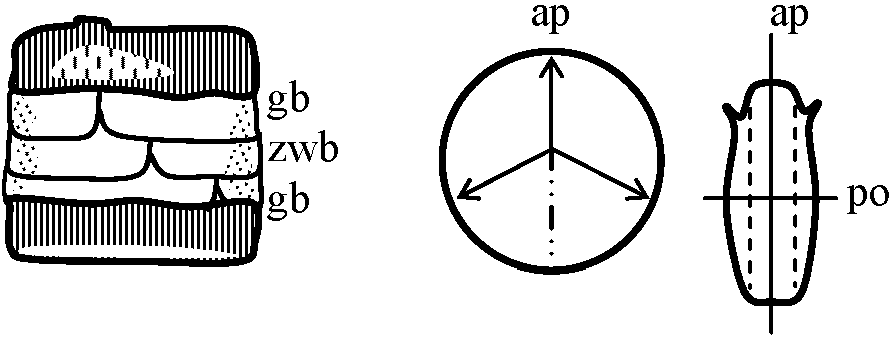
⊂ 黄色生物界 Chromista: 微小プランクトンから海藻を含む。水中 藻体: 運動性-不運動性 単細胞、群体、糸状体、嚢状体等、色緑、黄緑、黄褐色等 運動性単細胞のものは周皮で包まれる。運動性のものは遊走細胞1本、不等長2-3本の鞭毛を持つ 細胞壁を有するものは2片が重なり合った2重構造bivalent-structure多色素体: 円盤状disc、帯状band等 色素: chlorophyll a, c, eの他に carotene, xyanthophyllを多く含む 光合成産物: ロイコシン leucosin (chrysolaminarian)、油脂等 多くは細胞壁が二重構造をなし、光合成産物としてデンプンを形成しないことが他の群と大きく異なる 無性生殖: 2縦分裂、遊走子等。他に包嚢cystを形成。特殊なものは被嚢体stotospore、増大胞子auxospore形成 増大胞子 auxospore 有性生殖: 同型配偶子isogamete → 接合によるものと卵生殖oogamyがある淡水・汽水・海水中、湿土中に存在。浮遊性のものが増殖し、時に水の華water bloom形成。300属、10,000種 黄鞭藻鋼/ヒカリモ鋼 (Class Chrysophyceae)体制単純。有性生殖未詳。栄養体体制が分類第1基準黄緑藻鋼/不等毛鋼 (Class Xanthophyceae or Heterokontae)体制単純。有性生殖の知られているものは極わずか→ 体制発達段階に基準をおく分類大系が広く採用 珪藻鋼 (Class Bacillariophyceae), diatom(s)a) 藻体不動性単細胞 (一部群体)褐色-黄褐色細胞壁ペクチン質で、多く珪酸質を含み硬くなる = 被殻 frustule: thesilliceous wall → [上殻 epitheca + 下殻 hypotheca] 2枚の珪殻が重なり合う構造 → 様々な模様 = 分類鍵 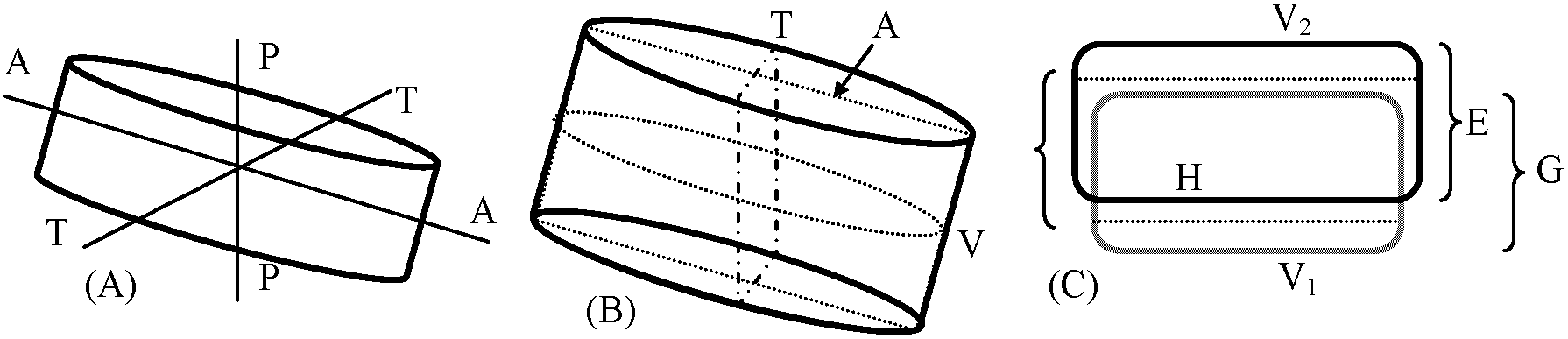 Fig. 4. A) 珪藻細胞の3軸。A-A: 頂軸 apical axis, T-T: 切頂軸 transapical axis, p-p: 貫殻軸 pervalval axis. B) 珪藻細胞の3断面。V: 殻面 valval plane, A: 頂軸面 apical plane, T: 切頂軸面 transapical plane. C) 珪藻細胞の被殻 frustule 模式図。V1V2: 上殻面と下殻面。G: 殻帯, E: 上殻, H: 下殻  Fig. 3. 珪藻細胞の被殻. Pinnularia viris. A. 帯面観 Overlapしている上殻と下殻。B. 殻面観。C. 切頂軸面(頂軸面に直角方向)。D. Rhizosolenia styliformis. s: 殻, gb: 殻帯, zwb: 中間帯 intercalary band, l: 貫殻軸(縦軸), q: 切頂軸(横軸), th: 分裂面, e: 上殻, h: 下殻, r: 背線 |
b) 色素体円盤状、帯状、星状等で数個ピレノイド持つものもある クロロフィルa, c、カロチンの他に各種キサントフィル 光合成産物 = 油脂 c) 生殖無性生殖: 2分裂。接合を繰り返したあと、特有の接合子(増大胞子auxospore)形成有性生殖: 精子と卵による。精子は褐藻に近い d) 生息地 habitat淡水、汽水に広く分布する主要プランクトンSubclass Centricae 中心亜鋼不等長2本の鞭毛を持った配偶体を形成中心から放射状の模様 Order DiscalesOrder SolenialesOrder BiddulphialesSubclass Pennatae 羽状亜鋼蓋殻面が楕円形、卵形、舟型、針型で、長軸に対しほぼ直角に線か点の列となる模様
縦溝(縦裂溝, 背線) raphe Order Araphidales 偽背線Order Raphidioidales 原始背線Order Monoraphidales 単背線Order Biraphidales 両背線ハプト植物門 Division Haptophyta= 黄藻植物門 Chrysophytaから独立Chl a, c。葉緑体は葉緑体包膜に包まれる(不等毛植物に似る)
鞭毛と異なり細胞表面が微細な鱗片に被われる
有機質の鱗片あるいは円石(石灰質主成分とする鱗片)を体表面に持つ Class Haptophyceae ハプトネマ= 特異な小器官 (共有派生形質 symapomorphy) – 分類群根拠周縁ER, peripheral endoplasmic reticulum: 門のもう1つの特徴。細胞膜直下で細胞全体を包む小胞体でハプトネマを構成する小胞体にまでつながる Order Isochrysidales イソクリシスOrder Prymnesiales プリムネシウム Chrysochromulina, Corymbellus, Phaeocystis, Platychrysis, Prymnesium Order Cocolithales (Coccosphaerales) 円石藻 細胞表面に円石(CaCO3の円盤型鱗片)構造を持つ Calcidiscus, Coccolithus, Syracosphaera, Umbilicosphaera Order Pavlovales パブロバ |
|
= 炎藻植物門: 系統群は大幅変更がなされた (ついていけない) 無性生殖は殆ど2縦分裂。有性生殖不明。淡水、汽水、海水に広分布。主要海産プランクトン Class Cryptophyceae 褐色鞭毛藻(褐鞭藻)鋼運動性1-2個の収縮胞あり色素体: 普通2個。帯状か円盤状 Order CyrptomonadalesCryptomonadcaceaeペリプラストperiplast: クリプトモナス類細胞質ラメラ直下にある隙間なく相互に連結したタンパク質板から構成されている物質 (おそらく細胞膜の周辺体) Nephroselmidaceae Order CryptocapsalesPhaeococcaceaeOrder CryptococcalesClass Dinophyceae 渦鞭藻鋼2本の鞭毛を持つ単細胞藻類 |
Subclass Desmophycidae (Desmokontae) 帯渦鞭藻亜鋼Order DesmomonadalesOrder ThecatalesOrder DinophysialesSubclass Dinophycidae (Dinokontae) 渦鞭藻亜鋼Order PeridinialesOrder DinocapsalesOrder RhizodinalesOrder DinococcalesOrder DinotrichalesClass Chloromonadophyceae 緑鞭藻鋼Order Chloromonadales褐虫藻 zooxanthella: 海産無脊椎動物と細胞内共生する単細胞渦鞭毛藻類
大きさ約10 μm |
|
= シアノバクテリア (藍藻細菌) 原生植物: 核、葉緑体、ミトコンドリア、液胞等細胞器官欠
藍藻植物 + 細菌植物 = 原核生物 procaryophyta ⇔ 真核生物 eucaryophyta 光合成: 特別な器官持たず多数チラコイドが単独に分散状態で存在。光化学系II関与色素はフィコビリン phycopilin 貯蔵物質: 主に藍藻デンプン cyanophycean starch。ステロール系物質なし 細胞壁主要構成成分: ムコペプチド 生殖
体分裂 binary division (二分裂藻 Schzophyceae), and fragmentation 体制体制分化には2つの傾向非糸状傾向 1) 単細胞単独遊泳型 → 2) 不定形群体遊泳型 → 3) 定型群体遊泳型 → 4) 単細胞定着型 → 5) 群体定着型 → 6) 偽組織定着型 糸状傾向 1) 単列糸状型 → 2) 偽分枝単列型 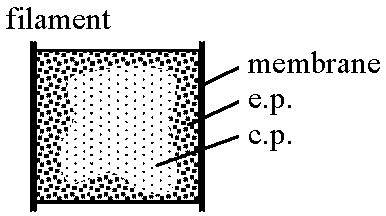 生活様式: 単細胞、群体、糸状体
生活様式: 単細胞、群体、糸状体
細胞糸 (毛状突起) trichome/鞘 sheath 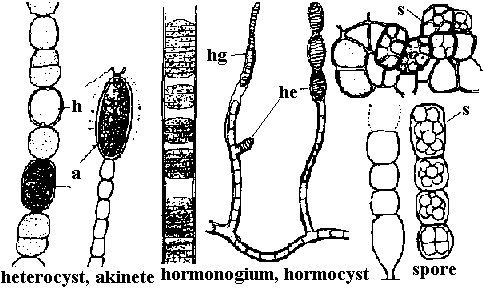 Fig. Reproductive organs in Cyanophyta. アキネート(耐久胞子, 胞子) akinete: トリコームを作る栄養細胞が貯蔵物質を蓄積し大型化し、厚膜を作り不利な環境に耐えるために変化した細胞 |
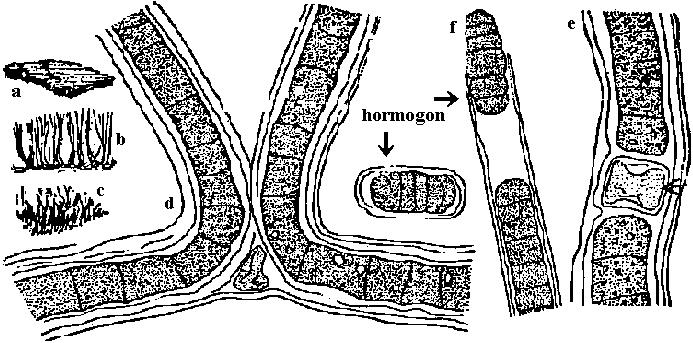 Scytonema coactile Montagne var. thermalis Geitler. h = 連鎖体 (hormogonium)
Scytonema coactile Montagne var. thermalis Geitler. h = 連鎖体 (hormogonium)(a-c藻塊全景, d, e 一部拡大, f先端) 藻塊オリーブ色-紫緑色、束をなすか密綿毛状に集まり広がる。偽分枝対をなし出るが稀に単出、鞘紫色。成層しない。11-18μm Φ, トリコームにくびれ無いが、時おり存し末端狭くなる。トリコーム細胞円筒形、9-11μm Φ, 7-20μm lでΦの1-2倍。異質細胞は円筒形で径はトリコーム細胞より僅かに広い。温泉産。宮城中山温泉・鬼首温泉。ジャワ Oscillatoria nigro-viridis Thwaites 藻塊黒緑色。トリコームはオリーブ緑色呈し幅7-11 μm、細胞間連接部括れる。先端へやや細くなり片側に曲がる。細胞短く幅の1/4-1/2倍、3-5 μm l。隔壁に沿い顆粒列。先端細胞は多少頭状に膨れ先端の膜はやや肥厚。汚水、岩壁、海岸杭等、温泉。九州、欧州、インド、北米 Oscillatoria chalybea Mertens var. chalybea: 藻塊暗緑色。トリコームは多少もつれる。真直から多少捩じれるか、不規則に螺旋状に巻く。細胞間連接部で少し括れる。幅8-13 μm、一端へ僅かに細くなる。先端部は鉤状に曲がる。細胞長は幅の1/3-1/2倍で3.6-8 μm。細胞間隔壁に沿い顆粒を有する場合がある。細胞内容は青緑色-暗色を呈し、微細顆粒を一面にしき、その間に少数の大形顆粒が散在する。先端細胞は鈍円で多少伸びる。静水、汚水、温泉。九州。世界各地 Aphanizomenon flos-aquae: 糸状体針状、束状藻塊。鞘見えない。浮遊。細胞形長円筒、4-6 μm Φ, 5-15 μm l。異常細胞円筒形6-8 μm, 35-80 μm l。道 Nostoc lincka: 藻体寒天質、布状不定に広がる。褐色-緑褐色。外皮なし。異常細胞ほぼ球形。道・本州
Suborder Stigonematinales Chroastipes linearis: 外部共生 Peliaina spp.: 内部共生 純粋培養 → 細胞内にシアノフィシン顆粒cyanophycin granules生産 ↔ 宿主細胞中では生産しない |
全て単子葉多年生植物
日本2-5科10属 (エングラー体系)ヒルムシロ (Potamogetonaceae) |
アマモ (Zosteraceae): スガモ (Phyllospadix iwatensis) トチカガミ (Hydrocharitaceae) |
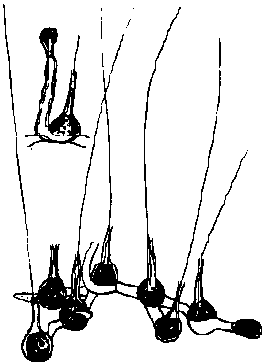
室蘭実習 (1983年)藻類生態調査: 実施日 1982年5月26日 場所 室蘭チャラツナイ浜目的: 室蘭における藻類の垂直分布及びそれに関する要因、更には、藻類の適応性を概観的に理解する。 方法: 岩盤上に生息する藻類分布状態を海面下から岩盤頂点まで幅15 cmのラインを求め、全長を15 cm間隔に区切る。つまり、15 cmずつのコードラート(Q)を行い、その相観を決定する。尚、Qでは海藻の種別の被度を採用した。 岩盤: 南向き斜面にA-Mの13 Q設置。Aが頂点。A-Dはほぼ平、J-Lはほぼ垂直、Mはオーバーハング。 結果: 岩の外観の所で使用した記号部各々をコードラートナンバーとした。被度はパーセント(%)を用い、数個体が若干見られたものはプラス(p)で表す0° 25° 50° 60° 95° 130° 総長 (cm) 65 40 35 15 35 15 コードラート A B C D E F G H I J K L M 傾斜 (°) 0 25 サイズ (cm) 15 15 15 20 15 15 10 15 20 15 15 20 15 Gloiopeltis furcata 65 (p) 50 5 60 5 (p) Pelvetia wrightii 5 80 30 90 10 100 100 75 75 30 (p) Analipus japonicus 2-3 (p) Chondrus yendoi 30 45 30 75 15 Ulva pertusa (p) 5 (p) (p) Gigartina pacifica 10 5 Monostroma undulatum (p) (p) Chondrus pinnulatus (p) 5 Laurencia nipponica (p) Rhodomela gracilis (p) |
更に、海面下に生息していたものの種名だけを記す(9 種) Kornmannia zostericola, Rhodoglossum japonicum, Phyllospadix iwatensis, Neodilsea yendoana, Alaria crassifolia, Costaria costata, Sargassum confusum, Tichocarpus crinitus, Odonthalis corymbitera 考察 エゾイシゲ、フクロフノリは岩の上部から中部にかけて広く分布している。このことは、この2 種は他の藻体より乾燥に耐えられる性質が強いことを示しており、その理由は、体中に気泡部を2 種とも有しているためらしい。実際に、この岩を見てみると、直接に波の影響を受けるのはH 部まで位と思われる。満潮時の波の影響というものも観察してみたい。 岩の中間部から、つまりH 部からはマツモ、イボノリ、クロバギンナンソウが出現する。H部を境に斜面は傾斜が変化する。このことは光を受ける要因と、水をかぶる要因の2 要因の変化が考えられる。これらの種の形態的特徴を考えると、体が多軸型で細胞組織と糸組織の分化があげられる。このことが乾燥への耐性の変化に関与していると思われる。 岩の下部、斜度が95o の面からイボノリ、シワヒトエグサ、ヒラコトジ、ウラソゾ、ホソバフジマツモが見られる。岩の斜度は以後90o を越すので、ここの生息種は、光吸収への適応が見られると思う。また波は干潮時以外は受けている、もしくは水面下と考えられるので、水への適応は十分と考えられる。 尚水面下にはモツキヒトエ、アカバギンナンソウ、(スガモ)、アカバ、チガイソ、スジメ、フシスジモク、カレキグサ、ハケサキノコギリヒバが観察され、これらは海水に常時つかっていないと生息は困難の様である。 以上のことから見て、この岩に置いての生態的分布は、上部から順に3 種に区分され、その要因としては、水への適応(乾燥への適応)が決定的なものになっていると考えられる。 |
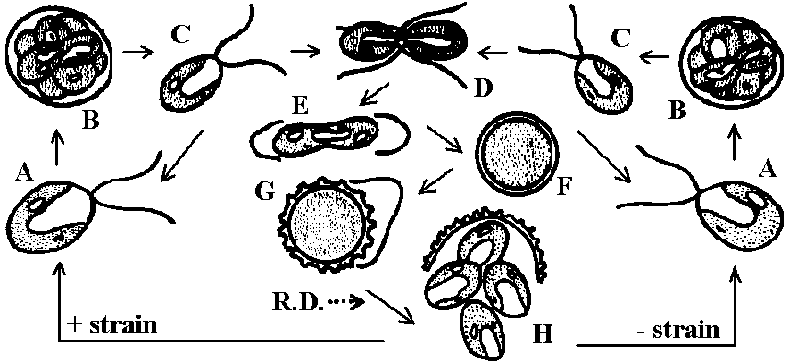
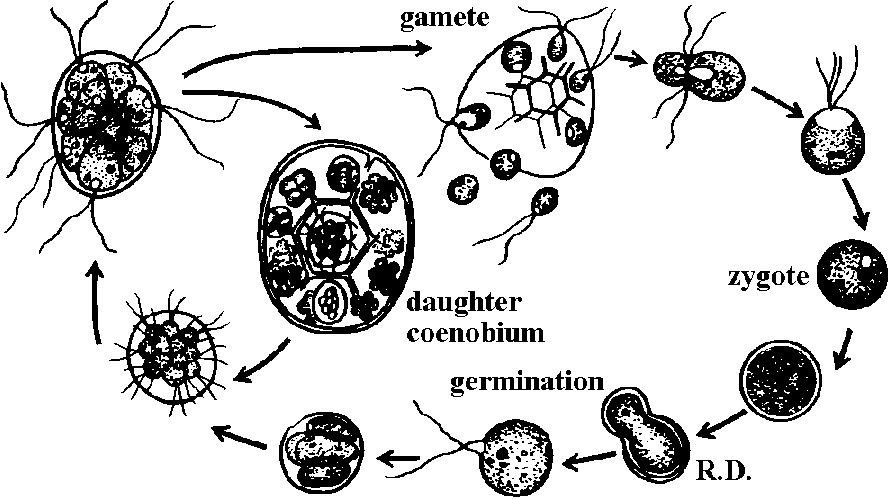

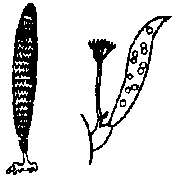
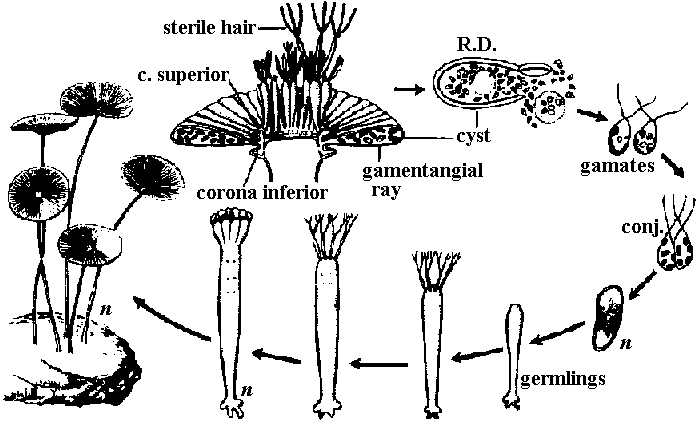
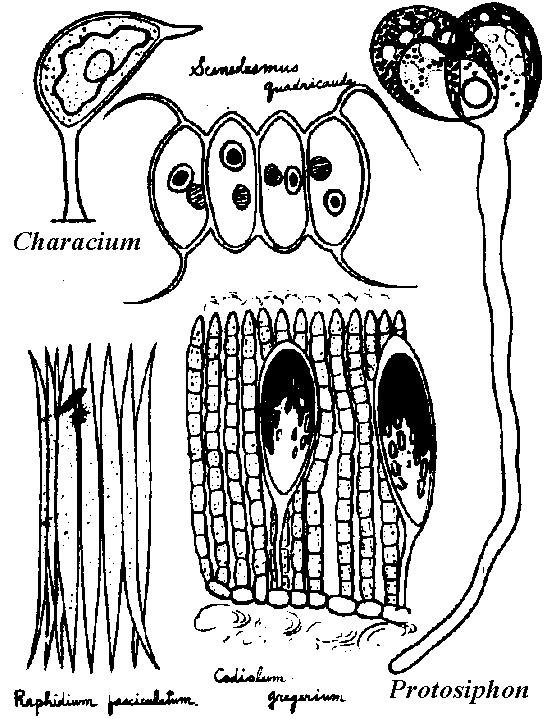
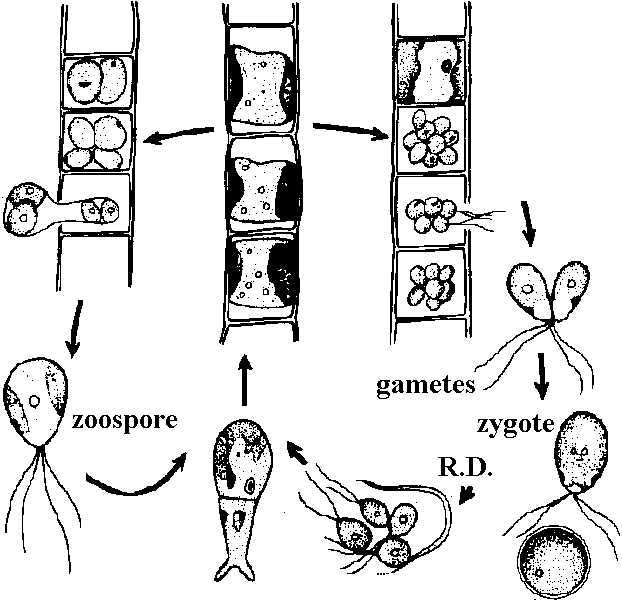
 → R.D. →
→ R.D. →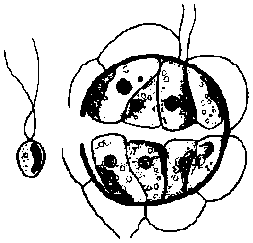 →
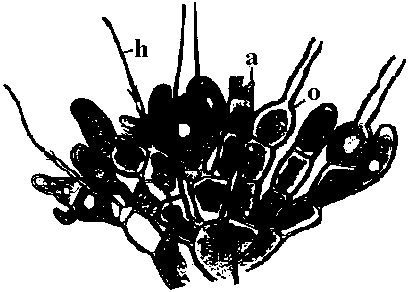
→ 
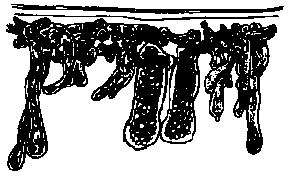


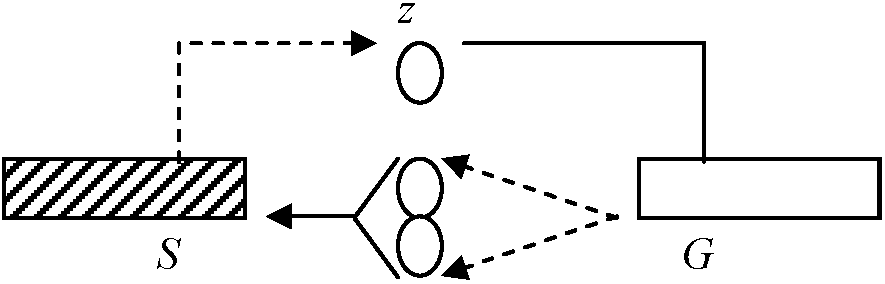 ムチモ型
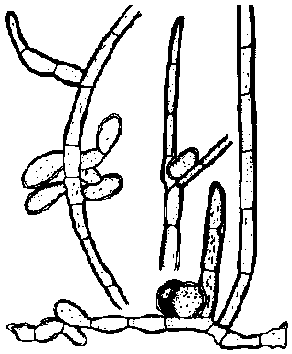
ムチモ型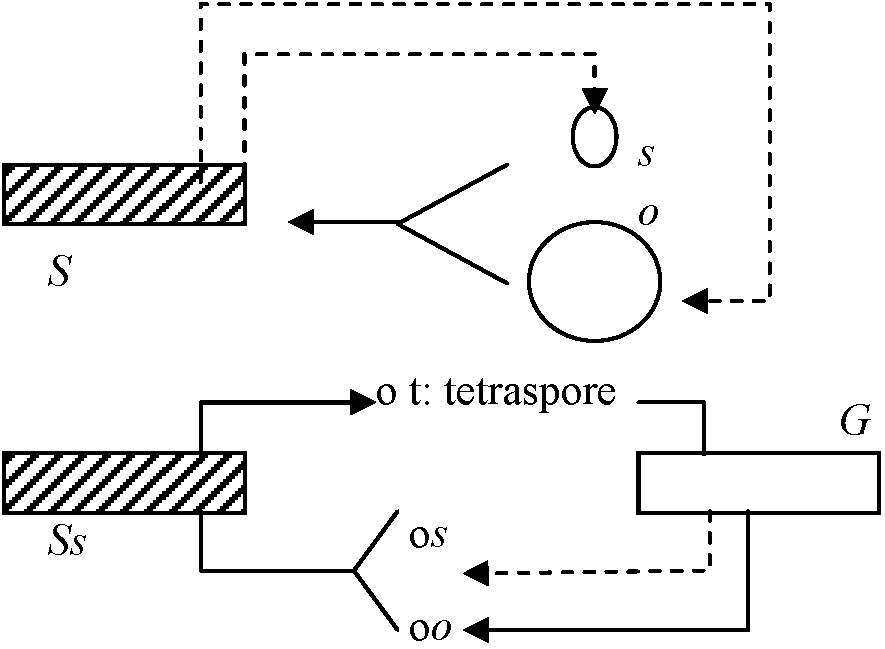 アミジグサ型
アミジグサ型