(2021年6月30日更新) [ 日本語 | English ]
HOME > 講義・実習・演習一覧 / 研究概要 > 小辞典 > 生活史
[ 生活型 ]
|
≈ 生活環 life cycle (生態学では) 個体の出生から死亡に至るまでの過程 → 生活様式に関する諸問題(環境・形態・生理等)を軸に生物の一生を表現 植物は、種子から発芽定着することにより生活史が始まると考えがちだが、生活史は生活環と言われるように1つのサイクル(life cycle)を有する。 Ex. 植物が最も移動できるときは花粉と種子の段階であり、動物でいえば種子は大人である。生理生態の解明を目的とするならば、次世代への適応戦略を頭に置くことが必要 世代 generation世代単位は、(無性繁殖を除けば)個体の有する遺伝子に変化が起こった時と考えるなら、種子(あるいはそれ以前の胚の発生段階)が新しい世代の始まり [ 蘚類 | 羊歯類 ]鶏が先か卵が先か: 生活環はどこから初めてもよい。ただし、個体の始まりを起点とするなら卵から |
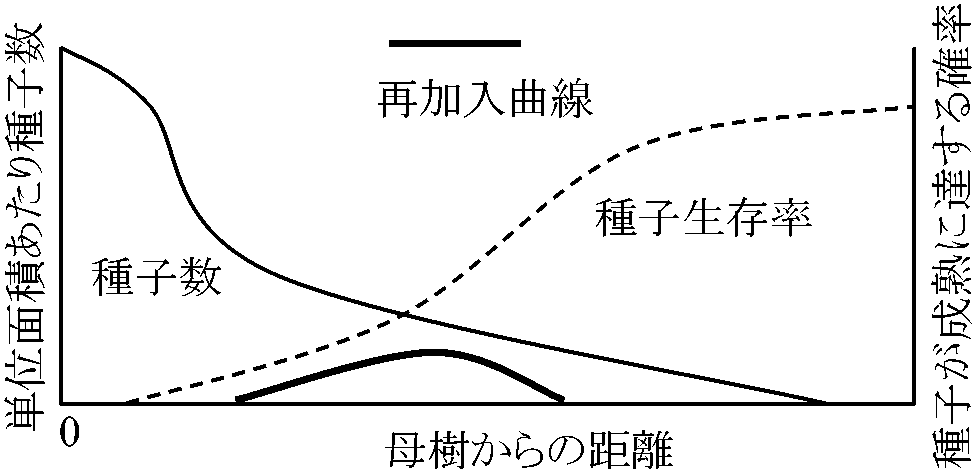
生活史段階実生 seedling発芽-成熟までの期間 → 植物生活系(一年生・多年生・木本等)により具体的期間は異なる繁殖までの成長様式が重要であり、関与する環境要因・競争関係の調査が繁殖成功測定にに必要 1932 Boysen-Jensen: 実生のジレンマ成長最適環境と発芽最適環境は異なることがある → 自分の生存により有利な生息地を選ぶならどちらかを犠牲にせねばならない(Aguiar et al. 1992) 1970 Janzen: 母樹からの距離と実生数(再加入数)の理論的モデル
|
グライムの三角形 (Grime's triangle)耐性 stress tolerance撹乱 disturbance 競争 competitive ability 一年生植物 annuals種子から種子までの生活を1年以内で終える草本植物 - したがって越年生は1年生に含まれる
夏型 1991 Barbieer et al.: 野生イネ2種16系統 乾燥地域において多年生 → 1年生進化 2005 Evans: Oenotheraの21種を種間比較降水量の少ない生息地に1年生草本が多い |
開花のタイミングQ. 繁殖活動と栄養器官成長との間で、光合成獲得物質をどのようなスケジュールで分配するのが最適かA._ dx/dt = (1 – u(t))g(x) dy/dt = u(t)g(x)
x: 栄養器官のサイズ 多年生植物 perennials2年以上にわたって生育する植物 = 一年生・二年生植物以外の全て宿根草 (≈ perennial)ストレスの高い時期(冬期や乾燥期)に、地上部を枯らし地下部で生存する戦略をとる多年草宿根: 多年生植物の地下部 |
|
季節変化に伴う動植物の行動・状態・形質等の変化
Ex. 発芽, 開葉, 開花, 紅葉, 落葉 光: 太陽光変化に影響される生理的・生態的過程 (Chazdon 1988)1) 時間スケール
Ex. 光周性 + 生物時計(体内時計 Ex. 日周期 diurnal, or サーカディアンリズム)
|
春植物 (spring ephemeral)= 早春季植物春先に植物群集上層部の葉が展開し光量が急に低下する前に葉を広げ、開花結実し、光量が低下時には地上部を枯らし休眠状態に入る (例, examples) 熱呼吸= 開花時寒冷対策
春、花芽成長時に数cmの雪を被るが、発熱で融雪し氷雪にトンネルを掘り、花軸が伸び、その上につく花芽が雪上に出て開花する 展葉落葉性植物の葉の開葉・落葉戦略は2タイプに分けられる(菊沢 1986)
一斉展葉タイプ: 極相林など安定な場所
個体小 → 遅くまで葉を展開 ⇒ 大きくなるにつれ次第に展葉早めるよう変化(予測) 花の寿命 longevity of individual flowers他殖率(c)の推定式(Primack 1995): 直接開花数が花の寿命に影響していることを仮定c = 1/(fp)
c: 他殖率 outcrossing rate
c = 1/(3 × 10) = 0.033 ≈ 3% |
[ 繁殖/生殖 ]
植物での進化仮説自家交配回避 - 近交弱勢回避花粉補償 捕食者回避 ディスプレー (雄花) 雌雄雌(♀): 大型で養分を多量に含む配偶子を作る個体 – 初期の発生を賄う雄(♂): 小型で運動力に優れた配偶子を作る個体 – 養分ほとんどなし 性型 sexual formCase. 植物雌雄同株(単性雌雄同株・雌雄異花同株・雌雄混同株・雑居花・雑性花) hermaphrodite and monoecy (synoecious): 雌性配偶子と雄性配偶子が1個体に作られる= 両性花と雌花又は雄花 male flower (staminate flower)が同一株につく
雄性先熟 protandry
雌雄同熟花 adichogamous flower 装飾花(不稔花) ornamental flower: 特に大型で美しい Ex. ガクアジサイ、ヤブデマリの装飾花 雌雄異花同株 monoecism, or monoecy: 単性花を形成 (♂/♀)単性花 unisexual flower: ♂f = 雄蕊、♀f = 雌蕊 → 一方のみ有する Ex. カボチャ、キュウリ、スイカ 雌雄同花序 androgynous: 花序に雄花・雌花が混在するもの 三性同株{(雄性雌性両全性同株) androgynomonoecious, trimonoecisous: (♂/♀/♂♀)雄性同株(雄性両全性同株・雄花両性花同株) andromonoecious: (♂/♂♀)
自家交配を避ける |
雌雄異株 dioecism, dioecy: 雌性配偶子個体(雌株) + 雄性配偶子個体(雄株)(♂)/(♀) Ex. ホウレンソウ、Morus, Ginkgo、ソテツ、スギゴケ、ゼニゴケ
三性異株(雄性雌性両全性異株) androgynodioecious, triecious: (♂)/(♀)/(♂♀)
雌株は花粉を作らない分、より多くの種子を生産できる (不完全雌雄異株 subdioecy, 雌雄混株のsubdioecious) → 多系 (雌雄混株・雑居性雌雄異株) polygamous, polygamny: 離性内の不完全雌雄異株のうち雄性異株・雌性異株を除いたもの
polygamomonoecious: 多性polyecy (adj. polyecious)が一部見られる不完全な雌雄同株 Case. 動物雌雄同体 hermaphrodite, or monoecy (monoecious): 1個体に卵巣と精巣を持ち、卵と精子を作る動物(普通卵成熟期と精子成熟期が異なり自家生殖起こらない) Ex. ミミズ、マイマイ、サナダムシ、ジストマ、ホヤ
常時雌雄同体 simultaneous hermaphrodite 性決定 sex determination[ 細胞遺伝学 ] 性染色体 sex chromosome環境 environment雌ホルモン, 酵素, 年齢, 水温等
|