(2013年4月27日更新) [ 日本語 | English ]
HOME > 講義・実習・演習一覧 / 研究概要 > 小辞典 > 垂直分布
|
≡ 標高勾配(elevational gradient)に沿った植生帯の発達 成因: 環境勾配 + (上部・下部との)種間競争 途中相では温度反応による植生分化が進まず分布帯細分化は困難なこと多 垂直分布 = 緯度 + 標高(気温減率) + 山体サイズ + 独立峰か否か + … Ex. 大雪山(N44°, 標高2290 m): 山体大きく高山帯は1600-1800 m以上 寸詰まり現象(≈ 山塊現象・山塊効果): 垂直分布の圧縮された状態 - 独立峰
独立峰 = 熱放散大 ↔ 山塊 = 熱放散小 |
垂直分布 (vertical distribution)高山帯 (alpine zone)森林限界 forest line 亜高山帯 (subalpine zone)山地帯 (montane zone) = 低山帯 山麓帯 (lowland zone) ≈ 丘陵帯 |
|
Def. 森林限界線より上の分布帯: 15 ≥ WI (最低気温 -30°C) (☛ ツンドラ)
年中降雪し万年雪も見られる お花畑: 森林限界より上部で高山植物が占める個所(地域) (明確な定義なし) 成育期間極めて短い + 乾燥・低温への適応 Def. 森林限界線 timber or forest line: 垂直分布で樹林帯上端(一般に明瞭)形成要因 = 地形・方位 → 気温 + 風向風速 + 積雪量等
Ex. 稜線部よりは風衝から守られる谷の方で森林限界上昇 独立峰山頂効果: 低標高でも山頂付近は森林低木化 ≠ 森林限界 土壌移動: 植物定着できず森林限界下げるEx. 富士山: 砂礫テフラ - 土壌移動 - 宝永山森林限界は太郎坊(1400 m) Def. 樹木限界 tree line: 木本植物定着できる限界風衝(偏形)樹: 高木が下枝広がり幹が旗状で芽風下に偏した形
+ 強風 (Ex. 森林限界付近、海岸)  高山風衝草原: 種数多。多積雪だとハイマツ群集等も形成され、それに伴い腐植質の蓄積もあり土壌発達
高山風衝草原: 種数多。多積雪だとハイマツ群集等も形成され、それに伴い腐植質の蓄積もあり土壌発達ミヤマシオガマ・チョウノスケソウ・イワオウギ・オヤマノエンドウ・ミヤマダイコンソウ・ヒゲハリスゲ・ミヤマノガリヤス等
岩礫地: 種子植物殆どみられない |
大雪高山植物群集: ヒース群系所生要素(マルバヤナギ・ミネズオウ・イワウメ・キバナシャクナゲ) + 岩礫地群系所生要素(チシマイワタデ・タルマイソウ・メアカンキンバイ) ハイマツ群落(低木林)森林限界指標
日本: 温度からは高山帯は全てハイマツ低木林 - 他要因で他群落形成 風上 = ハイマツ群落 ⇔ 風下 ≠ ハイマツ群落 日射量十分な安定斜面。冬期適度な積雪に覆われ芽保護される場所に発達ハイマツは寒冷期先駆種として進出し、針葉樹林発達と共に後退し、環境要因が針葉樹林を阻止する高山帯での極相と考えられる ハイマツ群落は北方に行くとササ的性格、つまり他の針葉樹林か広葉樹林の樹幹下に存在する低木とり、ハイマツと他の植物(群)との混成群集となる日本: 高山環境(風、乾燥、雪等)で各群落が分離 + 形態適応 modification + 種内分化 morphosis 各群落分布は様々な要因の総合的結果として決定 Ex. 日本でハイマツは一般に高所にあるが、低所における種間競争を避け高所に環境適応し分布したとも考えられる。しかし、低所でも、日高蛇紋岩地帯等では高所強酸土壌地域での生存が困難であるため低所でもハイマツ群落が観察される 背腹構造要因: 雪圧差本州: 風下側 = 吹き溜まり形成 → 積雪深2-5 m。ベタ雪で重い → 風下側にハイマツ生育できない 北海道: 軽雪で3 m積もることは少ない → ハイマツ発達可能 = 北海道でハイマツ群落は背腹構造示さない 高山植物 alpine plant日本: 4回にわたる氷河期(100万年-1万年前) = 寒冷地植物南下 ⇒ 気候温暖 = 北上 + 一部の植物は北上せず高山に移動 ⇒ 現在の高山植物環境(激変) = 強風 + 低温 + 強光 (UV含) + 積雪 + 低水分 + 低養分 ⇒
環境適応 Ex. 小型化(花は相対的に大)、地下茎・根発達 = 栄養貯蔵 巨大植物 giant plants一般に高山では標高上昇につれ植物サイズ小型化 → 逆の振る舞いEx. Giant Senecio, Puya |
|
= 針葉樹林 Ex. シラビソ・コメツガ・トウヒ 最低気温-20°C。降霜期間8-10ヶ月。土壌風化不十分。風・霧強い。日射量少 先駆種pioneerにはカラマツ・ダケカンバ等があり二次遷移初期はダケカンバであることが多い。ダケカンバは亜高山帯先駆樹種として遷移初期に優占する他の侵入種よりも樹齢が長いことが多い 縞枯れ wave regeneration風衝斜面: 縞枯れ現象やそれに近い一斉枯死の現象
大規模: 中部・大峰山系(シラビソ、オオシラビソ優占林のみ) - 南向斜面 森林限界直下、森林限界なければ山頂付近
土壌: 薄い (4-30 cm、10 cm程度の厚さが多) 偽高山帯 pseudo-alpine zone背腹構造: 日本海側 ↔ 太平洋側水平分布・垂直分布 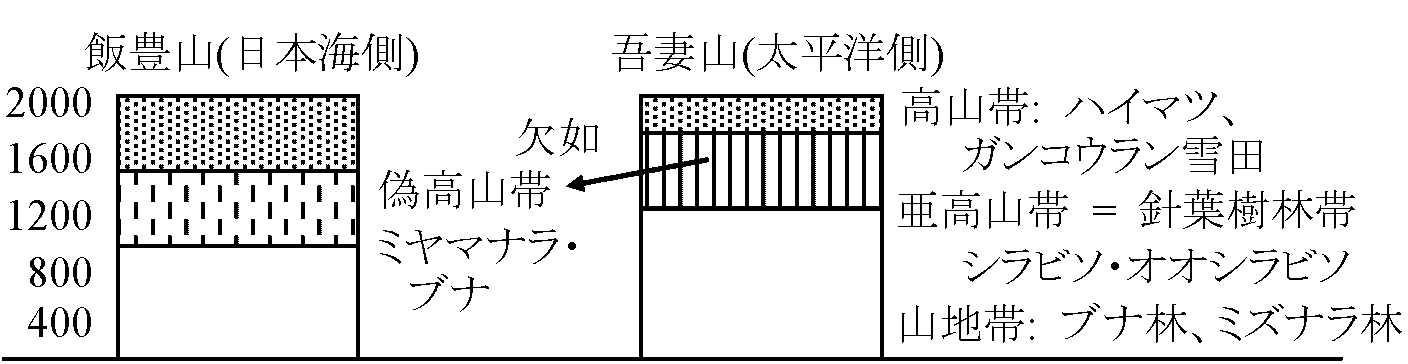 → 偽高山帯を発見命名 (四手井 1956) ニセコ・カリバ: ミヤマナラがある。北海道でも狩場山等ではブナの代りにダケカンバが生える 表。主な山岳における針葉樹林帯の有無。▲ 針葉樹林帯有、△ 無
日本海側 太平洋側
△ 岩木山 1625 (m) ▲ 八甲田山 1590
▲ 八幡平 1578
△ 鳥海山 2230 ▲ 栗駒山 1628
△ 月山 1979 △ 船形山 1500
△ 朝日山 1870 ▲ 蔵王 1841
△ 飯豊山 2105 ▲ 吾妻山 2024
要因: 地史的差(梶 1982): 第4紀第4氷期終了後の気候と植生成立の関係表. 第4氷期以降の気候変遷: 尾瀬ヶ原での花粉分析と14C年代測定から区分 *: 野尻湖
Nakamura Tsukada Nakamura Nakamura Yasuda
1952 1967* 1967 1972 1978, 1980
L 15000 -10000 12000-9000
RI = 寒冷 10500 -10000 -8000 -8500BP
RII = 温暖 9500 -4500 9000-4000 8000 RIIa -6500BP
(3000) 境界変更 RIIb
RIIIa 減温暖┓ 4500 -1500 3000-1500
RIIIb ┛ 1500BP-現在 1500-現在
|
氷期には現在より6-9°C温度低く、1000 m程度森林限界が現在より低かった 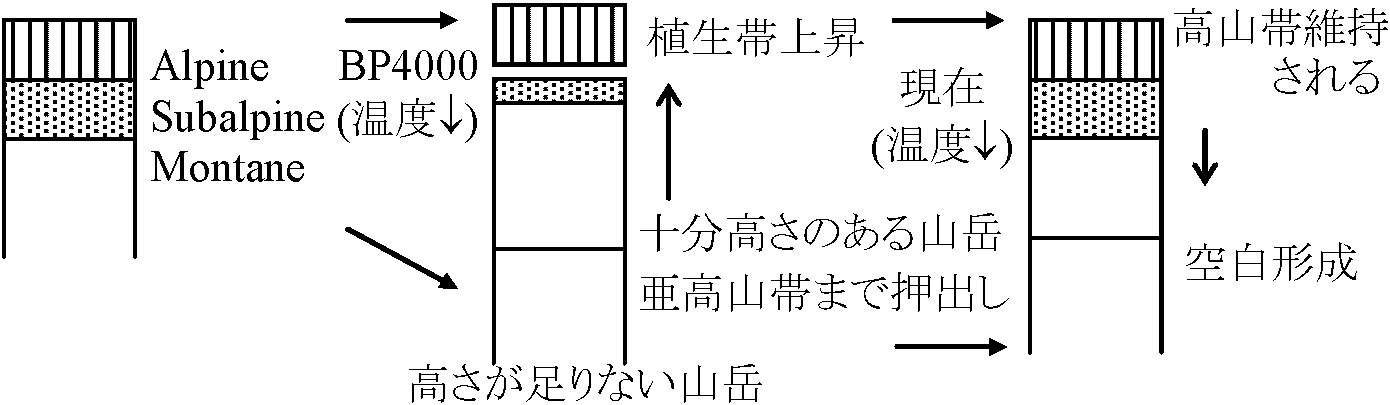
この空白を埋めるもの → 偽高山帯?
最終氷期針葉樹 = アカエゾマツ類、カラマツ類、チョウセンゴヨウ (鈴木・竹内1989) 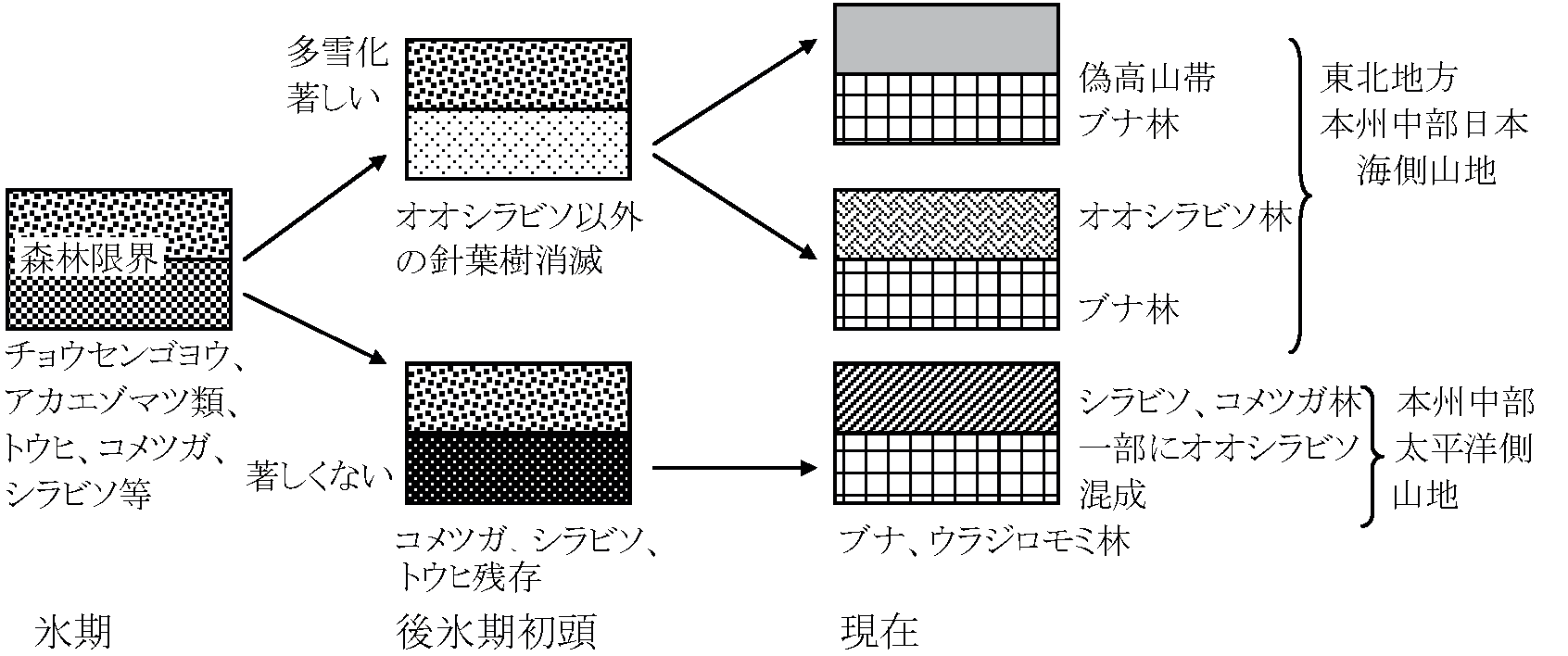
|
|
≈ 落葉広葉樹林(夏緑樹林) 最低気温 -10°C 降霜期間6ヶ月(上部): Fagus crenata・ミズナラ・シラカンバ 最低気温-5°C 降霜期間4ヶ月(下部): クリ・ケヤキ・クヌギ・コナラ 先駆的に林を作る → アカマツ・カラマツ・シラカンバ等 本州中部 ≈ 標高700-1700 m |
|
= 低地帯, 低山帯, 丘陵帯 ≈ 常緑広葉樹林(照葉樹林) 日本南部: シイ・タブノキ・カシ・クスノキ / ツバキ |
|
地史的には新山 + ハイマツ(Pinus pumila)がない! 樹木限界に匍匐カラマツ
ハイマツ: 新生土壌に侵入遅い → 匍匐型カラマツが森林限界構成(説) 垂直分布3776 m2400 m: 森林限界 _______↑ 亜高山帯 1600 m ↓ 山地帯 |
溶岩台地に植物が育ちはじめた痕跡生命力のない台地に最初に宿る植物たち溶岩に覆われて間もなくは、土壌が形成されていず、植物の姿を見ることができない。しかし、次第に地衣・コケ類が侵入し溶岩の表面を風化させる。さらに生きものの遺体が加わり、土壌が形成されていく。この場所の足もとでは、溶岩上に地衣類やコケに覆われ始めた頃の様子を見ることができる。写真: キゴケ / ハナゴケ / カリヤスモドキ, 花期は8-10月 溶岩流上に森が発達する過程での、このゾーンの段階:初期溶岩台地 → 地衣・蘚苔類の侵入 → 草本群落の発達 → 極相林 2018年12月9日 山梨県富士山科学研究所にて |
| 標高 (m) | 極相林 | 途中相林 | 草原 | 夏期平均気温 (°C) | |
| 4000 | -雪線- (気候的雪線) | 0.5 | |||
| 2500-3000 | ハイマツ | カラマツ | ウシノケグサ, コメススキ, イワノガリヤス, ナガハグサ, ササ |
構造土 高山草原・ハイマツ群集 |
+12 (年平均0) 氷河周辺気候 |
| 1500-2500 | シラビソ | ダケカンバ | -森林限界 (2500 m)- 針葉樹林 |
||
| 1000-1500 | ブナ | シラカンバ | シバ, ススキ, トダシバ, ネズミノオ, アズマネザサ |
落葉広葉樹林 | 森林気候 |
| 500-1000 | カシ | アカマツ | 常緑広葉樹林 | ||
| 0-500 | シイ | コナラ |
図. 極相林・途中相及び草原の垂直分布帯(蘚苔類・地衣類帯を除く)
|
2290 m ----- ① ハイマツ帯: 森林限界を超えた高山帯 コマクサ, ハイマツ 1500 m -----② 針葉樹林帯: 亜高山帯 ダケカンバ, エゾマツ, トドマツ, アカエゾマツ 800 m -----③ 山地帯: 落葉広葉樹林帯か針広混交林帯 (mixed forest) ミズナラ林, ミズナラ-トドマツ林等, 一部にトドマツ林とアカエゾマツ林 → 風穴 (wind cave) 図. 北海道大雪山における一般的な植生の垂直分布(模式) |
[ 噴火履歴 ] 羊蹄山 Mount Yotei |
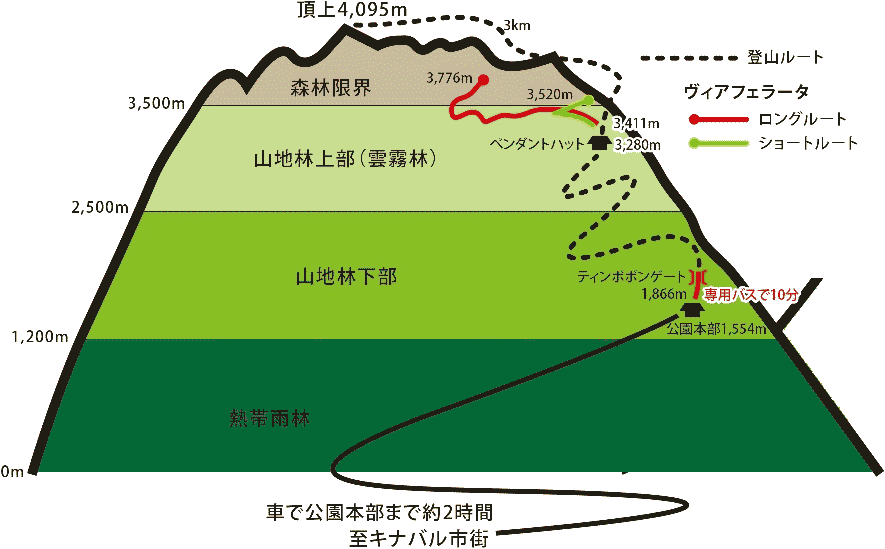 図. キナバル山(北緯6°)森林垂直分布 |