(2025年11月17日更新) [ 日本語 | English ]
HOME > 講義・実習・演習一覧 / 研究概要 > 小辞典 > 両生類・爬虫類
|
デボン紀末期出現 (多く絶滅 - 現世3目) 幼時: 水中で鰓呼吸 → 変態後: 肺呼吸 + 四肢形成 = 陸上生活可能 体表: 鱗・毛・羽はない + 産卵: 卵生か卵胎生 = 乾燥に弱い 心臓: 二心房一心室変温動物
無足目 Gymnophiona Müller 1832= アシナシイモリ/ハダカヘビ生息地: 森林内湿った地中 (一部水中性) 有尾目 Caudata Scopoli 1777Fam Hynobiidae Cope (サンショウウオ)Hynobius (サンショウウオ)H. abei Sato 1934 (アベサンショウウオ), RDB = EN
日本(石川県南西部-京都-福井)固有種 四国固有種 (過去H. boulengeri四国個体群とされる) H. retardatus Dunn 1923 (エゾサンショウウオ), RDB = LC道固有種, 2n = 40 (Syn. Satobius retardatus Adler et Zhao 1990) H. dunni Tago 1931 (オオイタサンショウウオ), RDB = VU日本(大分、熊本、宮崎一部)固有種、天然記念物 (大分市霊山個体群) H. tosashimizuensis Sugawara et al. 2018 (トサシミズサンショウウオ): 独立 H. boulengeri Thompson 1912 (オオダイガハラサンショウウオ), RDB = EN
天然記念物 (奈良、三重) 島後島固有種、天然記念物 (隠岐の島町) H. nebulosus Temminck et Schlegel 1838 (カスミサンショウウオ)
日本広域種 →
H. vandenburghi (ヤマトサンショウウオ): 近畿東部-東海南部 日本(東北・関東・中部)固有種 H. tsuensis Abe 1922 (ツシマサンショウウオ), RDB = NT対馬固有種 H. tokyoensis (トウキョウサンショウウオ) Nakamura et Ueno 1963
RDB = VU, 日本(関東-福島)固有種 2022 北部個体群 = イワキサンショウウオ(新種) H. lichenatus Boulenger 1883 (トウホクサンショウウオ), RDB = NT
日本(北関東-東北)固有種 天然記念物 (白馬村) ⊃ H. tenuis Matsui 1987 (ヤマサンショウウオ) H. kimurae Dunn, 1923 (ヒダサンショウウオ), RDB = NT日本(関東以西)固有種 H. naevius Temminck et Schlegel 1838 (ブチサンショウウオ), RDB = EN九州北西部固有種 H. stejnegeri Dunn 1923 (コガタブチサンショウウオ), Syn. H. yatsuiH. ikioi Matsui, Nishikawa et Tominaga, 2017 (ベッコウサンショウウオ)
RDB = VU (Syn. H. stejnegeri, auct non Dunn 能登半島北部・富山県固有種: 天然記念物 (羽咋市) Salamandrella Dybowski 1870 (キタサンショウウオ)S. keyserlingii Dybowski 1870 (キタサンショウウオ), RDB = NT 分布: カザフスタン-モンゴル-ロシア-日本(釧路湿原)-国後・色丹 天然記念物 (1975 釧路市、1992 標茶町) S. tridactyla (エンカイシュウキタサンショウウオ): 分枝系統から独立 Onychodactylus (ハコネサンショウウオ)O. japonicus Houttuyn 1782 (ハコネサンショウウオ): 分子系統 - 分離・独立
O. nipponoborealis (キタオウシュウサンショウウオ): 東北北部個体群 中国山地・岡山県: O. japonicusと同所的分布 Fam Cryptobranchidae Fitzinger 1826 (オオサンショウウオ)Andrias japonicus Temminck 1837, syn. Megalobatrachus japonicus Temminck オオサンショウウオ(ハンザキ)
ハンザキ: 半分に裂かれても再生する(という言い伝え) … Fam Salamandridae Goldfuss 1820 (イモリ, newt)Cynops (イモリ): 分布: 中国-日本C. ensicauda Hallowell 1861 (シリケンイモリ): 日本(奄美群島・沖縄諸島)
C. e. ensicauda Hallowell 1861 (アマミシリケンイモリ)
分布: 日本固有種 (道は内地からの移入)
南西諸島固有種 (台湾?) - 天然記念物 (1978 沖縄県、2003 鹿児島県) |

無尾目 Anura Fischer von Waldheim 1831ヒキガエル Fam Bufonidae Gray 1825アマガエル Fam Hylidae Rafinesque 1815鼻腔から目を通り鼓膜にかけ黒い模様 (アオガエルになし)アカガエル Fam Ranidae Rafinesque 1814アカガエル Rana• エゾアカガエル pirica Matsui 1991: ♂ 46-55 mm, ♀ 54-72 mm 樺太-北海道-国後島・択捉島 繁殖期: 雪解け直後, 平地-山地の水辺 → 離れる(草地・林床)  • トノサマガエル nigromaculata: ♂ 55-80 mm / ♀60-90 mm
• トノサマガエル nigromaculata: ♂ 55-80 mm / ♀60-90 mm水掻 = 後足のみ 体色 = 雌雄異: ♂全体緑-金色基調に背中央1本緑-黄色縦縞 ↔ ♀黒-灰暗色で背中中央縦縞太白 繁殖期 = 2月(水田張ると開始) 背中にミミズ状細長隆起散在し、背側線(鼻先-瞼)を通り尻まで続く線模様太く盛り上がる
トウキョウダルマガエル: ミミズ状隆起弱く残るが、背中に黒色の丸みを帯びた斑紋散在 日中も活動し昆虫類・クモ・ザリガニ等捕食。水田や周囲緑地依存 分布: 本州(関東-仙台地域と信濃川流域除く)、四国、九州、中国大陸 • トウキョウダルマガエル porosa porosa: ♂35-75 mm / ♀45-85 mm 背中央に1本の緑色-黄色縦縞持ち、縦縞途中細くなり途切れることある 体色 = 緑-金-灰暗色: 雌雄差余りない 平地・低丘陵地の水田・小川・池、湿地等。生態トノサマとほぼ同 分布: 関東-仙台、信濃川流域 = トノサマ空白地に分布、新潟長野等の一部地域で分布重なる → ×関東 = 関東「殿様ガエル」は本種 足トノサマより短 • ダルマガエル porosa brevipoda: ♂35-60 mm / ♀40-70 mm トウキョウダルマガエル亜種(命名規約) → 絶滅危惧は本亜種 小型で手足短 形態 = 地域集団 - 典型は背中中央に縦縞なく背中や足に丸い黒斑紋散在 餌: 小型昆虫類やクモ、ヤスデ等 雌一部は年2-3回産卵 平地水田、低湿地 → 生活史全てを水田依存する地域多 → 水田開発に弱 分布: 本州(伊那谷-東海-近畿-中国地方瀬戸内側)、香川県 繁殖期 = 4-7月 • ウシガエル catesbeiana: > 15 cm (日本最大) 水掻 = 後足水掻大, 体色 = 緑, 繁殖期 = 4-6月 体型頑丈 鼓膜大で直径目より大 背中黒斑点散在 平地水田や溜池、湖、ダム湖、緩流河川等。基本的に水辺生活(汚水可) 乱食: 幼体-成体まで他カエル捕食 繁殖期: 雄池中で縄張作り「ブオンブオン」と鳴く 分布 ≈ 全国 → 合州国原産帰化(食用に大正時代以降輸入) → 野生化 • ツチガエル (イボガエル) rugosa: ♂30-40 mm / ♀35-50 mm 全身縦長のイボ。青臭 指先端やや細い。背中中央に背中線ことがある 水掻 = 後足 吸盤なし 体色 = 土色 繁殖期 = 6(-8)月 平地丘陵地小川・水田付近。水辺から余り離れない 比較的清流な少流ある水辺好み関東都市部衰退 雄: 岸や水中でギューギューと鳴く 雌: 卵塊分け水草に付着産卵 一部は幼生年内変態せず越冬 分布: 北海道一部(人為)、本州、四国、九州 Fam Dicroglossidae Anderson 1871 (ヌマガエル)ヌマガエル Fejervarya Gravenhorst 1829• ヌマガエル (沼蛙) F. kawamurai Djong, Matsui, Kuramoto, Nishioka et Sumida 2011, Indian rice frog (Syn. Rana limnocharis) 30-70 mm (地域差大) 手足短い肥満型, 後足水掻ツチより小 背中に縦長線状隆起疎(ツチほどイボない) → 奄美・沖縄諸島産個体は背中線ある個体多。腹真白(ツチ腹には斑模様) 吸盤 = なし 体色 = 茶褐-黄土 水田付近等。概ね水辺生活 雄: 水田畦や草陰(道路側溝でも)に隠れるように鳴く 雌: 数回に分け水草等に付着させるよう産卵 繁殖期 = 5-8月(本州) < 奄美・沖縄 分布: 静岡県以西本州、四国、九州、奄美大島諸島、台湾・中国、東南アジア 日本産カエルで唯一、本州と奄美・沖縄諸島に分布 → 人為的分布拡大 • sakishimensis (サキシマヌマガエル): 先島諸島Fam Rhacophoridae Hoffman 1932 (アオガエル)上顎に歯Subfam Buergeriinae (カジカガエル) Buergeria Tschudi 1838 (カジカガエル) Subfam Rhacophorinae (アオガエル) Zhangixalus Li, Jiang, Ren and Jiang 2019
Z. schlegelii (シュレーゲルアオガエル) Günther 1858: 固有(本州・四国・九州・五島列島) Fam Microhylidae Günther 1858 (ヒメアマガエル)Microhyla okinavensis Stejneger 1901 (ヒメアマガエル): 北海道から遥か南 |
|
石炭紀出現 - ペルム紀繁栄
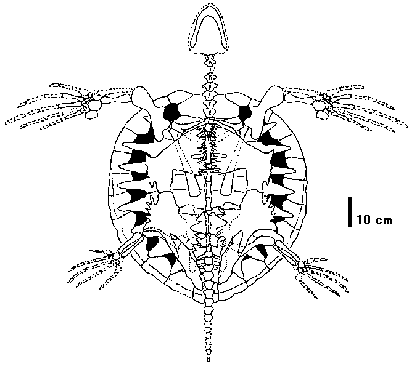
肉食爬虫類 → 草食爬虫類 染色体WW, WZ型 [恐竜群!]角質の体鱗に覆われた変温動物 無弓亜綱 Subclass AnapsidaTestudines カメ目 turtles, tortoises= Chelonia日本6科13種(含外来種(発見例少除), 亜種分類諸説) 甲羅 背甲 carapace: 背面にある甲羅
項甲板 cervical: 背甲頭部側先端にある左右の縁甲板を繋ぐ甲板
喉甲板 gular: 腹甲のうち一番頭部に近い位置にある左右に1対の甲板 爬虫綱 Class Reptilia= 爬虫類 reptiles無弓亜綱 Subclass Anapsidaカメ目 Testudines (= Chelonia) turtles, tortoises双弓亜綱 Subclass Diapsida
有鱗目 Squamata: ジュラ紀以降トカゲ亜目 Lacertilla (Sauria) lizardsヤモリ gegcko
前肛孔 = 総排泄孔(肛門)前小窪み モササウルス類 (Mosasauroidea): 白亜紀後期繁栄した海棲爬虫類 - 現代トカゲ近縁(見解によるが化石種) |
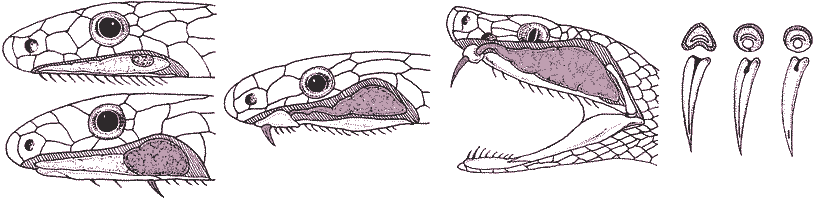
ヘビ亜目 Ophidia, snakes ピット pit: 特定ヘビで赤外線(温熱)を感知する感覚器官 北海道: クサリヘビ Fam Viperidae (ニホン)マムシ Gloydius blomhoffii Boie 1826 Protobothrops Hoge et Romano-Hoge 1983) ハブ: 全て夜行性 分布: 南西諸島で飛び石状に不連続 - 規則性が見つからない ハブ P. flavoviridis Hallowell 1861: 南西諸島固有、有毒(血清有)Syn. (旧) Trimeresurus flavoviridis Boulenger 1890 (波布/飯匙倩) トカラハブ P. tokarensis Nagai 1928: トカラ列島(宝島 小宝島)、低毒(死者0)Syn. (旧) T. f. tokarensis Nagai 1928 サキシマハブ P. elegans Gray 1849: 八重山諸島、低毒(重症例有)
Syn. (旧) T. elegans Gray 1849 (ハブと交雑可) Syn. (旧) T. okinavensis Boulenger 1892 (方言 マムシ) 不連続分布説明仮説地質説: 燐鉱石・硫黄噴出する島に非分布 (否定) 沖縄: ハブ追い出しに硫黄燃やす ↔ 硫黄産する子宝島にトカラハブ 温度説: 温度勾配に沿い分布決定 (否定)南西諸島では温度は南北方向に連続変化 - 規則性有 降水説: 降水量の多い島に生息 (否定)ハブは湿った場所(= 降水量の多い島)を好む 例外有 Ex. 降水量 宮古島(ハブ不在) > 石垣島(在) 標高説(半沢 1935): 石灰岩 = 標高低い島 (< 170 m) vs 火成岩 = 高い
氷河期: 琉球列島は大陸と陸続 - ハブ類分布拡大 琉球石灰岩堆積部 = 琉球期に海面下にあった
→ 海水面下がると低い島も現れるがハブは移入できない アオダイショウ Elaphe climacophora Boie 1826 シマヘビ Elaphe quadrivirgata Boie 1826 ジムグリ Elaphe conspicillata Boie 1826 シロマダラ Dinodon orientale Hilgendorf 1880: 夜行性 - 遭遇稀 鰐(ワニ)目 Crocodilia, crocodile三畳紀以降アリゲータ alligator: 閉口時に下顎4番目の歯見えない クロコダイル crocodile: 閉口時に下顎4番目の歯見える – 鼻吻部尖る。獰猛 |
|
Def. 主に人間に対し直接的・間接的に危害を加える可能性のある生物 (日本自然保護協会 1994) 対応(安全対策): 注意、救急措置、薬物、器具、保険 1. 予防 = 何もしない (a. 近寄らない(遠くから避ける) b. 触れない(食べない)
防衛できない生物には遭遇しない予防が第一
接触したら: 対応は種により全く異なるの = 予備知識重要
準備: 器具・薬品 ⇒ 応急処置 + 治療
生物界
※ 注意点: 場所(生息地)と時間(季節・時刻) ☛ 哺乳類 哺乳類生食は全ての種で寄生虫感染の危険大
ガス壊疽 gas gangrene: 血液供給不足等により壊死した組織が発酵菌に侵され腐敗 (自分の調査に直接関係ないのでメモ程度に) 海産動物危険: 外部有毒器官(最も危険) + 内部有毒物貯蔵器官 + 機械的障害☛ 魚類 ☛ アカエイ Dasyatis akajei (Müller et Hente), ray
尾根元付近に有毒刺(アカエイ科の種は全て刺を持つ) - 絶対に触らない 他にガンギエイ、シビレエイ等々
刺されたら早急に医師の治療 テンジクダツ T. acus melanotus (Breeker): 2種とも主に暖海(× 道)
堅く鋭い吻部による負傷(照明に突進すること多)
毒棘(= 早急に医師の治療)
潮間帯下部(浅瀬)
浅海の岩礁・サンゴ礁
鰭にある刺は全て毒腺が付随
沿岸岩礁に多
鋭歯 = 深くかまれると刺傷 (魚類ウミヘビ = 無毒)
毒ハゼ(シガテラ毒)、マングローブ林に多
潮間帯の岩礁・サンゴ礁
潮間帯岩礫地・サンゴ礁タイドプール等
殻長1 mに達する - 水中で挟まれると水死することもある
暖海、サンゴ礁(インド洋-オーストラリアまで)
速やかに病院搬送 (2時間で死亡例あり)
マダコ: 顎歯については同じ ラッパウニ Toxopneustes pileolus (Lamarck): 2種とも熱帯系 (× 道)
棘に有毒物質 - 触らない、踏まない
サンゴ礁: イシサンゴ類ポリプ捕食 - 分布概ね一致
+ ウモレオウギガニ、ヒラアシオウギガニ 食しない - 食べたら直ちに病院 (死亡率高)
岩の多い海岸(野外での実害はない)
絶対に挟まれない(挟まれたら外傷対応)、中毒死報告あり
無毒、鋏による外傷 - 一般外傷の手当(十分な消毒)
刺咬傷: 水洗
ジンマシン様皮膚炎時: 抗ヒスタミン剤含むステロイド軟膏塗布
小甲殻類: イソイツブムシ、フナムシ
剛毛(毒毛): 触れると激しい疼痛 - 決して擦らない
セロテープや消毒用アルコールでそっと拭き毒毛除去
ゴカイ、イソメ類: 強力な顎歯 - 咬まれたらステロイド軟膏 |
カツオノエボシ Physalia physalis utriculus La Martinniere: 暖海 (× 道)
上部に直径10 cm程度の気泡体(目印)
特別な治療法なし (死亡例あり) - アンモニア水、重曹水等を塗る
小形(≈ 3 cm): 8-9月頃日本沿岸漂着
処置はカツオノエボシと同じ
+ 刺胞毒激: アマクサクラゲ、アカクラゲ、ヒクラゲ、キタカギノテクラゲ、イラモ
ヒドロ虫の群体: 刺胞持つ - 人体に触れると射出 → 刺痛 (溺死例あり)
アンモニア水や重曹水を塗る アナサンゴモドキ(沖縄) ウデナガウンバチ Megalactis griffithsi Saville-Kent
刺胞毒: 触手
アンモニア水塗る、抗ヒスタミン剤塗布
ハナブサイソギンチャク 節足動物・環形動物有毒・感染症媒介等の危険生物多傷害様式
毒針 Ex. サソリ(尾端に毒嚢)、ハチ、ハリアリ
海外貿易のため港湾地区では予期せぬ被害に注意
応急手当: 圧迫止血、抗生物質内服
毒成分 = ヒスタミン、活性ペプチド (3亜種: アオズムカデ、アカズムカデ)
後腹部尾状6節 - 第6節に毒嚢と鉤状の毒針
毒針なし(サソリとの区別点) - 尾端から酢酸臭の液(触れない)
秋に雌はススキ葉を巻き巣とする - 咬傷: ステロイド軟膏塗布、水湿布
山林・原野のササ地に多 - 調査・登山で注意
口器が残ったら外科
ヤマトダニ、タカサゴキララマダニ
ネズミ駆除
幼虫がヒトに吸着しリンパ液を吸う ☛ ドクガ Euproctis subflava Bremer ☛ チャドクガ Euproctis pseudoconspersa Strand マツカレハ Dendrolimus spectabilis Butler
幼虫: 年一回発生(4-6月) = 毒針毛 + 繭外側にも毒針毛 (成虫は無害)
幼虫: 全身に毒刺を持つ肉角 (成虫・繭は無害)
幼虫: 黒褐色瘤根部が丘状に膨らみ毒針毛有する (成虫・繭は無害)
触れない → 抗生物質含むステロイド軟膏塗布
カンタリジン含む虫体液 - 発赤、腫腸 → 水泡
北海道: アオカミキリモドキ、ツグロカミキリモドキ、ツマグロkァミキリモドキ、キクビカミキリモドキ (× 道: シリナガカミキリモドキ、ハイイロカミキリモドキ)
触れない - カンタリジン含む虫体液 (手当: アオカミキリモドキに準ずる)
ヒメツチハンミョウ: (× 道)
キイロゲンセイ: 花上、灯火(夜)
水田、畑、池塘周辺、河岸。道分布
→ 抗生物質含むステロイド軟膏塗布
アシグロアリガタハネカクシ、エゾアリガタハネカクシ、アリガタハネカクシ ☛ 双翅目: カ(蚊) マツモムシ (半翅目) Notonecta triguttata Motschulsky: 道分布
刺傷: 掴むと刺す = 掴まない |
爬虫類蛇毒 (venom)捕食方法進化: 二次的に防御機構とし機能(種毎異なる Ex. 神経障害も様々)
↔ 両生類: 被食回避手段としての防御物質
安全: 1. 慌てない、2. やたらに縛らない、3. やたらに切らない
噛まれた患者: 1. 休ませる 2. 安心させる(数時間後も血清有効) 3. 噛まれた局部動かさない 4. 全身症状注意 5. 早く医療施設に連れて行く ワニ(×日本)・トカゲ(日本産はまったく無害) カメ有毒亀 (×日本): タイマイ = 餌からくる毒(タイマイ自体は無毒) - 摂食注意 |
スッポン Trionyx sinensis: 水底が泥となる沼・河川 (× 道)
咬むのは早く力は強い → 裂傷 (毒はない) - 通常の外傷処置
餌となるカエルの分布するところ (× 道)
大人しい - こちらから手を出さない限り危害はない
エラブウミヘビ Laticauda semifasciata: 海岸近い海 - 南西諸島 マムシ Agkistrodon blomhoffi blomhoffi: 頭扁平で正三角形風 - 他種と区別
50 cm以上離れていればほぼ安全
ヒメハブ・トカラハブ: 死亡例なし 両生類日本には特に危険な種は分布せず一部に毒分布腺・筋肉中に毒 - 取扱い注意
Ex. ヒキガエル類、アマガエル類、イモリ
フグ毒と同じ物質 - 接触後よく手を洗う(手に取った手で目・口触らない)
他にナガレヒキガエル・オオヒキガエル (× 道, 3種とも) |