(2026年3月3日更新) [ 日本語 | English ]
HOME > 講義・実習・演習一覧 / 研究概要 > 小辞典 > 地質年代 > 化石
化石化 (fossilization)陸水域や海水域で起こること多断片: 葉、茎、幹、果実、種子、花(花粉)、球果、胞子 葉、幹、胞子は化石化しやすい 化石堆積年代 - 古くなるほど化石の持つ情報が減るのが普通化石fossil再構成rework: 1. 不完全性, 2. 二面性, 3. 異地性化石産物で現在残っているもの
軟体部/硬体部。又、その物質交代が見られるものもある 連続切片は有効な方法 Ex. 甲冑魚: 神経配列観察可(発電機?) ナノ化石 nanofossil: これらの微細構造単位で用いられる化石 化石生痕学(古生痕学) palaeoichnology or ichnolithology生痕化石 tracks or trackwayに関する研究1912 Abel: Lebensspuren (life spur) = 過去の生痕 足跡 footprints、這跡 trails、巣穴 blurrows、穿孔跡 borings、糞石 coprolite、苦悶死、病型、共生、胃石 gastrolite 足痕学 pedalogy: 足跡 Ex. 化石人類学糞形学 coprology: 糞(排泄物)研究 - 食性・腸の形と機能 古病理学 palaeopathology: 化石に残る病型・奇形 (Moodie 1923) 化石残存形態
分類: 化石上の個体と種完全個体復元 - 1個体認められ、1種とし記載
器官属 organ genus: 器官ごとに与えた属名
≠ 生物学的分類群 taxon あるカテゴリー(類型化、階層)に従い、人為性ないもの |
成長: 化石上は硬組織のできる度合いgrowth rateを主にさす 環境(温度、水分等)を反映する成長様式存在 → 古生態・古生物地理 古生物: 分類結果 ⇒ numeical taxonomy ≈ classical taxonomyとなること多視点を変えると、古生物では化石の不完全性が問題
Ex. Desmostylus mirabiris Nagao (desmo- 束の, stylus 歯): 樺太第3紀中新世地層中 → 古生物学: 雌雄片方発見 → 一方でよいがholotype記載必要 組織保存硬組織保存 → 軟組織Rugosa: カルサイト形成。絶滅種 化石の問題 = 後世変形(化石形成後の化石変形) → 化学作用、熱作用る変形は多く考慮した再現必要 Goreau: サンゴの代謝 → 日中はCaCO3吸収盛んだが夜は静か
現在のサンゴの1年にできる縞数 ≈ 360 形態研究法
化石発見→
産状記録→採集→運搬→
試料調整→登録→
写真
計測→
観察記録→個体発生→
古生態・古気候→
古地理分布→ 系統 |
古環境復元 paleoenvironment生活圏 biotope (≠ ビオトープ): 化石となる古生物の本来の生息地 遺骸圏 thanatope: 遺骸の分布範囲
異置性 allochthonous: 生活圏 ≠ 遺骸圏
特徴(形状・大きさ)等から種と適応度・環境再現 - 行動 fossil behavior 深度: 広深性 eurybathyal ↔ 狭深性 stenobathyal 鹹(塩)度: 広鹹(塩)性 ↔ 狭鹹(塩)性 食物: 広食性 euryphagic ↔ 狭食性 stenophagic 温度: 広温性 eurythermal ↔ 狭温性 stenothermal 表. 鹹度による水域区分
淡水 低(貧)鹹水 中鹹水 高(多)鹹水 海水 技術巨視的: 材、動物骨等 + 微視的: 花粉、細胞 |
A. 石化標本: 有機物が鉱物に置き換わり化石化された標本 接線法によるセクション: 横断面 - 葉形, 縦断面 - 内部形 = 組織調査 → レプリカ法 replica method (剥ぎ取り法 peel method) 葉化石: Case. 組織(表皮、柵状組織、海綿状組織、維管束等)を保存 角皮 cuticle: 酸化剤処理で残る(他は分解) - 気孔含む表皮構造 シダ植物では表皮も分解(= 角皮が残るのは種子植物) ⇒ シダ種子植物とシダ植物の判別に使える B. 岩石標本: (地質学の)研究教育のために集められた様々な種類の岩石a) 浸漬法 maceration method 細胞や組織間の結合を機械的、超音波、酸、アルカリ、キレート、酵素等により分離する。化石では、表面構造を観察しやすくするため用いる b) レプリカ法 peel method 表面磨き希酸エッチング → 表面に凹凸 → レプリカreplica i) 炭化葉: 石から剥がす peel → シュルツ溶液 Schultz solution 浸漬 → 水洗 → 10% KOH → 水洗 → 表皮残る → プレパレーション preparation ii) 炭化材: HF浸漬 → KOH 2-3日浸漬 → ミクロトーム薄片作成 → プレパレーション C. 印象標本 impression: 生物の形状や構造が岩石に残されたもの内部形態観察不可能 → 外部形態観察(植物: 外部形態で脈系詳細観察可) コロジウム法: EtOH + コロジウム → impressionを写し取る |
古地質時代菌類(古菌類)研究 → 菌類起源・進化解明 (☛ 菌類)
化石菌類研究法1. 産地産状産地: 菌類多くは植物体内外付随 ≈ 植物化石産地(= 良質葉・材化石)= 河口泥質物・花粉含む地層等 + 石炭(= 植物化石)
大形サルノコシカケ、泥質物濾過により得る硬質の子嚢菌子実体等は植物体から離れ産出多 Ex. 生物体集積沈澱物からなる先カンブリア時代チャートに混在する菌糸状化石は藍藻と区別困難 Ex. 菌類胞子や菌糸と見誤りやすい微化石に、糸状繊毛虫類、シオグサ類、接合藻類化石がある 2. 採集法植物化石に付随する菌類化石採集法に準ずる = 植物化石含む堆積岩をツルハシ等で掘り取る植物化石痛めないよう採集 → 植物化石含むと期待される石はトンカチで割る 植物化石部位は割れやすく、菌類化石付随箇所は見当がつかないことが多く、痛めないよう採集し持ち帰る 3. 標本作成法と保存法菌類化石の状態に応じ作成法を選択採集資料: 多くは湿り、新聞紙や布に包み放置 - (現世)カビ生え化石菌類と見誤られ十分注意 泥中糞化石等: 乾くとヒビ入り原形失う - 採集後早く観察用プレパラート作成 → 作成標本は、データ付けプレパラート整理箱や紙箱、ガラス瓶等に保存 a. 菌類化石付随する葉化石(葉コンプレッション化石)からの標本作成 母岩から剥ぎ取れる場合 ピンセット等用い簡単に剥取れる カセイソーダ(NaOH)に浸漬し葉化石を母岩から遊離 + 同時に脱色
剥取った葉化石はポリビニール・ラクトフェノールを十分量用い封入 |
コロジオン被膜を葉化石表面に作り、溶媒蒸散後、慎重に剥しバイオライトで封入しプレパラート作成 b. 岩石中菌類化石: ダイヤモンドカッターと研磨装置を用い薄片作り、カナダバルサム等で封入しプレパラート作成 c. 泥質物中菌類化石: 篩を用い粒状物をふるい分け、その粒状物の中から菌類化石を選び出し78-80%C2H5OHや4-5%ホルマリンに浸漬保存 菌類化石菌類起源有力証拠提供する例稀(先カンブリア紀水中生物化石中未発見)
化石発見困難: 多くの菌類は肉眼観察困難 + 大型キノコは腐り易く化石化しにくい 先カンブリア紀: 細菌や藍藻化石に混じる2種菌糸化石(最古菌類化石) 菌類は20億年以上昔の先カンブリア紀に水生菌とし出現? デボン紀中期泥炭層: 接合菌亜門接合菌綱アツギケカビ科(?)菌類化石
デボン紀に菌類は陸上植物出現に付随し陸上に出現 北米石炭層: サルノコシカケの1種の化石発見 産出状況: 種類により菌類化石は沢山産出 → 葉表面や内部に生える子嚢菌類や不完全菌類多
大形キノコ化石: 多くはサルノコシカケ類。植物化石含む堆積岩(途中段階含む)中産出。サルノコシカケ以外にハラタケ類や腹菌類(ツチグリ)等があるが日本からは未報告 |
[ 生命の起源 ]
化石による証拠1965-70: 先カンブリア紀化石が低変成岩から多出
先カンブリア紀の岩は殆ど変成を受けた塩基性岩
藻類葉状体様 like-algal leaf = 藍藻 b) Bacteria chain spot: indirect proof and doubtful c) 石墨 graphite: denatured formation d) Chemical fossils カンブリア紀: 無脊椎動物多出 → O2の変化が原因と考えられる 1) 先カンブリア紀初期 early pre-cambrian
化石は(南)アフリカから産出。全てが球状藻類 チャート中から微生物(球状 10-100 μm) b) Fig tree group (3.1 × 109 yr): スワジランド Swaziland system
Onverwach groupの上層 ストロマトライトstromatoliteと呼ぶ石灰岩(ドロマイト)中より胞子と思われる化石と藍藻化石確認 ⇒ 光合成生物が表れている2) 先カンブリア紀中期 middle pre-cambrian a) Witwatersrand super group (S-group) (2.2-2.2 × 109 yr): 南アフリカ 帯状-紐状フィラメント |
b) Vallen group (2.0 × 109 yr)
硫黄バクテリア(?) – 多種化 (12種8属) c) Gunflint (iron) formation (1.9 × 109 yr): カナダ、シューペリオール湖
採取した鉄分豊富なチャートから植物化石多 – 最低藍藻6種と細菌2種 球状フィラメント 3) 先カンブリア紀後期a) Beck spring Dolmite (1.3 × 109 yr): 南カルフォルニア 藍藻とフィラメント 5-10 μm球状体 – 被核細胞 b) Skilogalee Dolmate (1.0 × 109 yr): 南・中央オーストラリア球状かフィラメント(緑藻化石 – 染色体持つ = Eukaryotes化石) c) Bitter spring formation (0.9 × 109 yr)球状かフィラメント – 細胞分裂中と思われる化石が混じる カンブリア爆発 the Cambrian explosion (or radiation)バージェス頁岩 Burgess shaleスティーブン累層の一部を構成する5億3千万年前(中期カンブリア紀)堆積層妙ちくりんな奴ら weird wonder (Gould 1989) 軟体性化石の保存状態良い
鱗甲虫 Halkieriid: カンブリア紀(絶滅) - 環形・腕足・軟体動物等の起源? 視覚器官を持つ → 食物連鎖(食う-食われるの関係)始まる 後に南アフリカ、シベリア西部等で類似した動物群発見エディアカラ動物群 Ediacara (or Ediacaran) fauna≈ エディアカラ生物群 Ediacara biota全球凍結解除後に突然現れる = 5億6千万年前(原生代最後期)化石群構成種の殆ど軟体性(刺胞動物?) = ベンドビオンタ 骨格ない軟体性動物の化石化は珍しい - 泥流等により生き埋め?
体制はコラーゲンにより維持 - 大量の酸素必要 ⇒ 全球凍結解除と光合成 → O2濃度上昇急激 → 大進化(仮説) Phylum Arcaeocyatha 古盃類先カンブリアン(絶滅)。海綿質(珊瑚礁)。日本はカンブリアン地層欠 |
|
先カンブリア紀: 生物全て海産 カンブリア紀: 藻類 → 半海水・湖・淡水に進出 珪藻・シャジク藻を除き藻類は出そろう 藻類化石出現 = 3.2-3.4 × 109 yr (問題: 陸上植物の起源?)= どのように淡水に適応したのか? |
陸棲藻: 緑藻 - 葉緑体含む
Ex. Fritshiella: 生活様式形態は蘚苔類近縁 vs 生殖細胞大きく異なる 陸上植物 = 中心柱 stele → 輸送器官 transportive tissue 水, 蒸散 evaporation, 土壌栄養 soil nutrients |
☛ 代: 古生代
カンブリア紀 Cambrian 541-485 Mya三葉虫類 [温暖化]
Archaeocyathus (古杯類), Redlichia (三葉虫), Paradoxides (三葉虫) オスドビス紀 Ordovician 485-444 Mya知られる最古の陸上植物: オルドビス中期(微化石) 470 Mya= クリプトスポア cryptospores - コケ植物に近い原始的形態
陸上植物(胚植物)胞子由来 (まだ遡るだろう) シルル(シルリア)紀 Silurian 444-419 Mya日本: 秩父古生層(シルル紀) + 北上山地古生層 ≈ 最古 →
サンゴ類 Ex. ハチノスサンゴ Favosites、クサリサンゴ Halysites デボン紀 Devonian 419-359 MyaHalysites, Leptanea, Megalodon, Phacops, Cephalaspis, Ichtyostega魚の時代 + サンゴ類カブトウオ出現 堅頭類出 現最古の森林 [温暖 → 寒冷化] 初期: 古生マツバラン(後期絶滅)、古生リンボク、古生トクサ程度 - 単純 Hyenia(古生トクサ): 15 cm h. Protolepidodendron (ヒカゲノカズラ類) Calamophyton: 0.8-4 m h 有節茎(トクサ的)、多条中心柱(シダ的)大型化: Hyenia → Calamophyton → Calamites 中期-石炭紀初期(期間4千万年): 陸上植物多種多様に分化
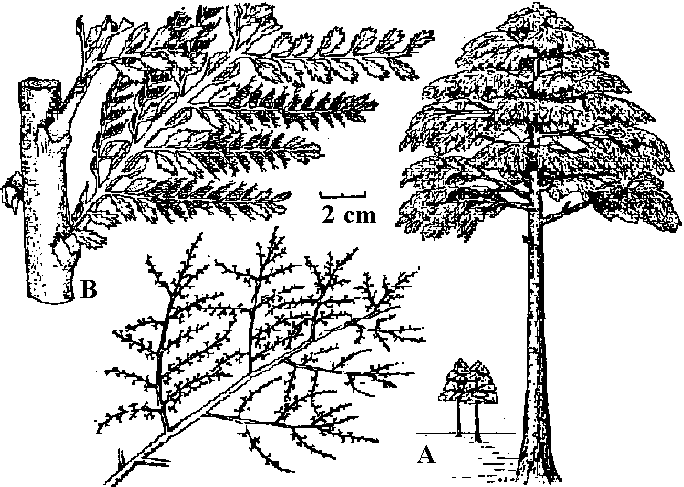
古生裸子植物、シダ種子植物出現 > 385 Mya 種皮発達 = 環境変化(乾燥等)耐性↑ + 捕食↓ + 休眠 1950 橘行一: Leptophloeum 岩手県上部デボン系(古生リンボク)フロラ (石炭紀-二畳紀)シダ種子植物の時代石炭紀 Carboniferous 367-289 Mya初期・前期(下部) = 陸上(石炭形成) ⇔ 後期(上部) = 海洋
隠花植物全盛 (Cycadales, Ginkgoales, Coniferales出現 → Permian繁栄) → 石炭70-80%この時期に形成 (☛ 炭球) Equisetopsida (s.s.) トクサ類(有節植物)Archaeocalamites: 石炭紀初期Calamites ロボク(蘆木): 植物全体と樹幹の化石の属名
最大高 20 m。茎中央に維管束、外側に細胞増やし成長 |
Lycopsida ヒカゲノカズラ類担根体 rhizophore: 有 = 二叉分枝することより分かるLepidodendraceae リンボク*: 石炭紀-二畳紀。湿地林要素。肥大成長活発 Lepidodendron リンボク(鱗木): 植物全体と樹幹の化石の属名
= 茎, 30 m h, 1-2 m d, 葉枕leaf cushion螺旋形
異型胞子(大胞子 600 × 16 spores, 小胞子 20 × 1000 spores) Triletes = 胞子
Lepidodendronに似るが、分枝0-2回程度 葉枕六角形、横長菱形-円形 二畳紀(ペルム紀) Permian 299-252 Myaシダ植物森林衰退 - 乾燥が原因? ☛ カタイシアフロラ三畳紀 Triassic 252-201 Myaジュラ紀 Jurassic 201-145 Mya白亜紀 Cretaceous 145-66 Mya |
ライニーフロラ(植物群) Rhynie flora1913 Mackie SJ発見: 当初デボン紀中期と推定 - デボン紀前期(4億年前)
ライニー(Scotland)付近の厚さ約2.5 mのチャート層 Rhynie chert 古生代植物群の変遷石炭紀 = レピドデンドロプシス植物群 + アンガラ植物群石炭紀終-ペルム紀 = ゴンドワナ + アンガラ + ユーラメニア + カタイシア ユーラメリカフロラ(欧米植物群) Euramerican floraデボン紀 (カレドニア造山運動: シルル紀終期-デボン紀初)
湿地帯発達 大葉類: Lonchopteris, Alethopteris, Supaia, Sphenopteris Archaeopteris: 最古の木本植物 (∵ 真正中心柱) アンガラフロラ Angara, Siberia or Kuznetsk flora石炭紀-二畳紀: クズネツク盆地(露)-シベリア-沿海州気候大変化が種分化と分布に大きな影響 (寒冷 ≈ ゴンドワナフロラ)
大葉系植物: 単葉はみない 小葉類: Noeggerathiopsis 有節類: Sphenophyllum, Paraschizoneura カタイシアフロラ Cathaysia or Gigantopteris flora古生代(石炭紀-二畳紀): カタイシア大陸 = 現中国大陸が中心1927 Halle: 山西省太原炭田
石炭紀(3 Bya): 海 ⇒ 二畳紀: 陸地上昇 ⇒ 三畳紀(2 Bya): 大陸 単葉の起源 ☛ 成長遅滞原則 (産出地: 中国、朝鮮、北米) 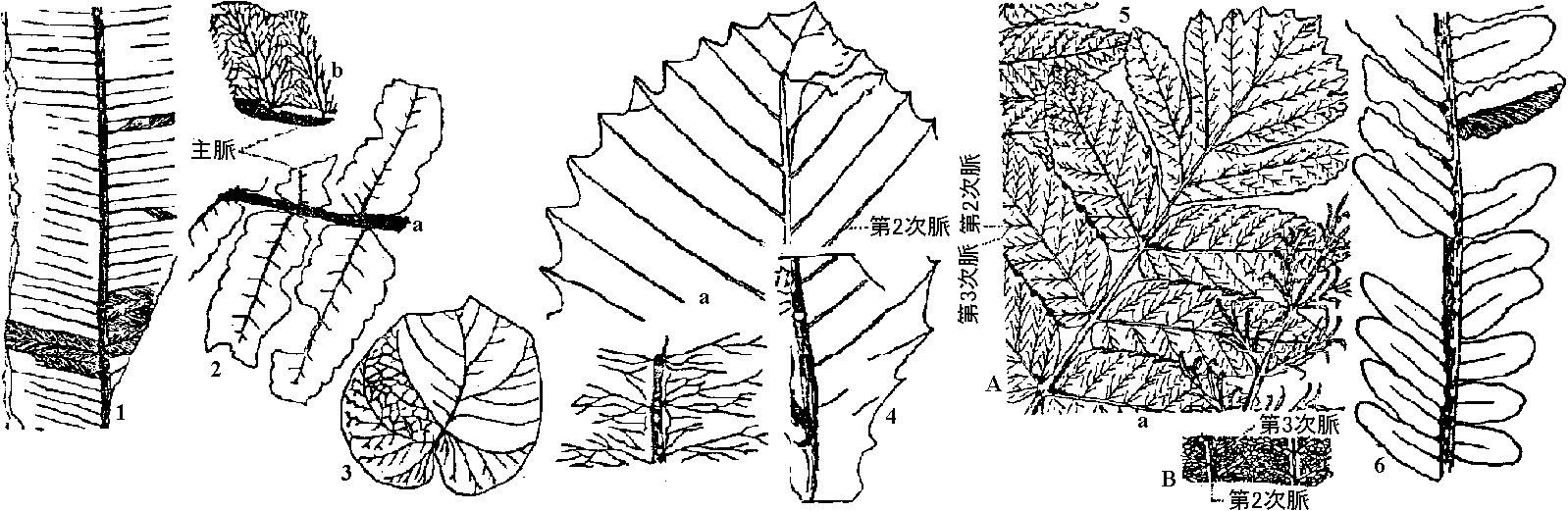 |
Fig. 1. Cathaysiopteris whitei (山西, デボン紀上部), 2. Gigantopteris largreli (1同), 3. G. cordata (福建, 二畳紀上部), 4. G. nicotinaefolia (朝鮮平安道, 二畳紀上部), 5. 4の復旧, 6. Neuropteridium polymorphum (山西, 二畳紀上部)
中生代以降この地域は乾燥化進む ゴンドワナフロラ(植物群) Gondwana or Glossopteris flora石炭紀末: 氷河 → フロラ一変 (熱帯性複葉植物消滅)二畳紀上部 upper Permian の氷河跡(氷堆積・擦痕)から記録 - 冷涼 石炭紀後期: ゴンドワナ大陸とみなせる地域のみから産出 ≈ Glossopteris相/植物群: 舌状単葉・網状脈 - 全体の9割占める
+ 大葉類: Palaeovittaria, Gondwanidium , ○ ゴンドワナ植物群 Gondwana flora")
Fig. 下部二畳紀植物分布。□ アンガラ植物群 Angara flora, [●欧州 Europe, △ 北米 North America] = 欧米植物群 Euramerica flora, ▲ カタイシア植物群 Cathaysia flora (東部アメリカを含む), ○ ゴンドワナ植物群 Gondwana flora 石炭紀上部からTriassic下部にかけては化石が、時間的にも空間的にも連続し産出する |
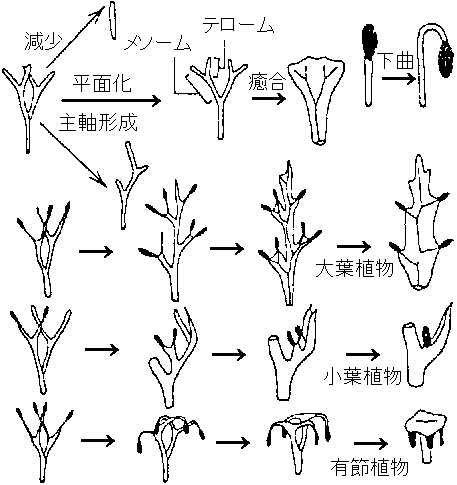
解決したい疑問1. 水中と陸上で進化の速度が違う(時代が新しいほど早い)2. 進化は一方向に起こる Ex. 単純 → 複雑 3. 進化の要因と機構 4. (維管束)植物の起源は単元(単系統)か多元か 巨大化石(macro-fossil)と微視化石(micro-fossil)がある 4.75 Bya (シルル紀): 上陸 - オゾン層形成により紫外線弱まる 蘚苔類 → 体制: 幹 - リグニン蓄積 → 維管束発達 1) 巨大化石 macro-fossil (蘚苔類 vs 羊歯類 ☛ 退行進化)2) 微視化石 micro-fossil
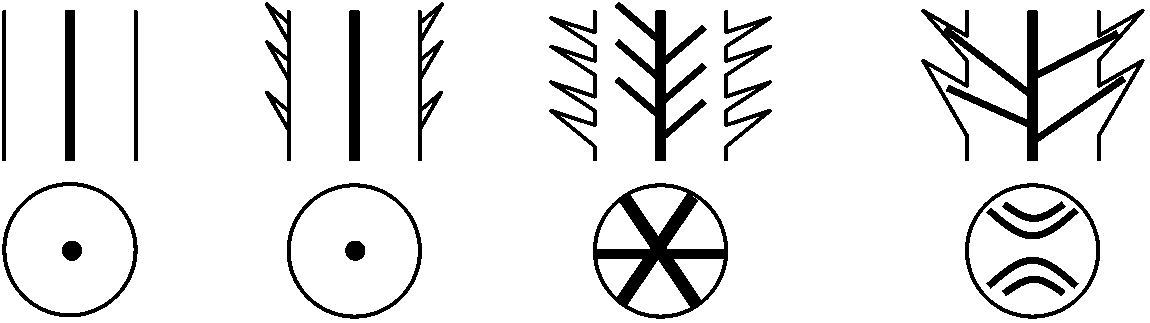
Pollination – 花粉学 palynology: pollen or spore, epidermis 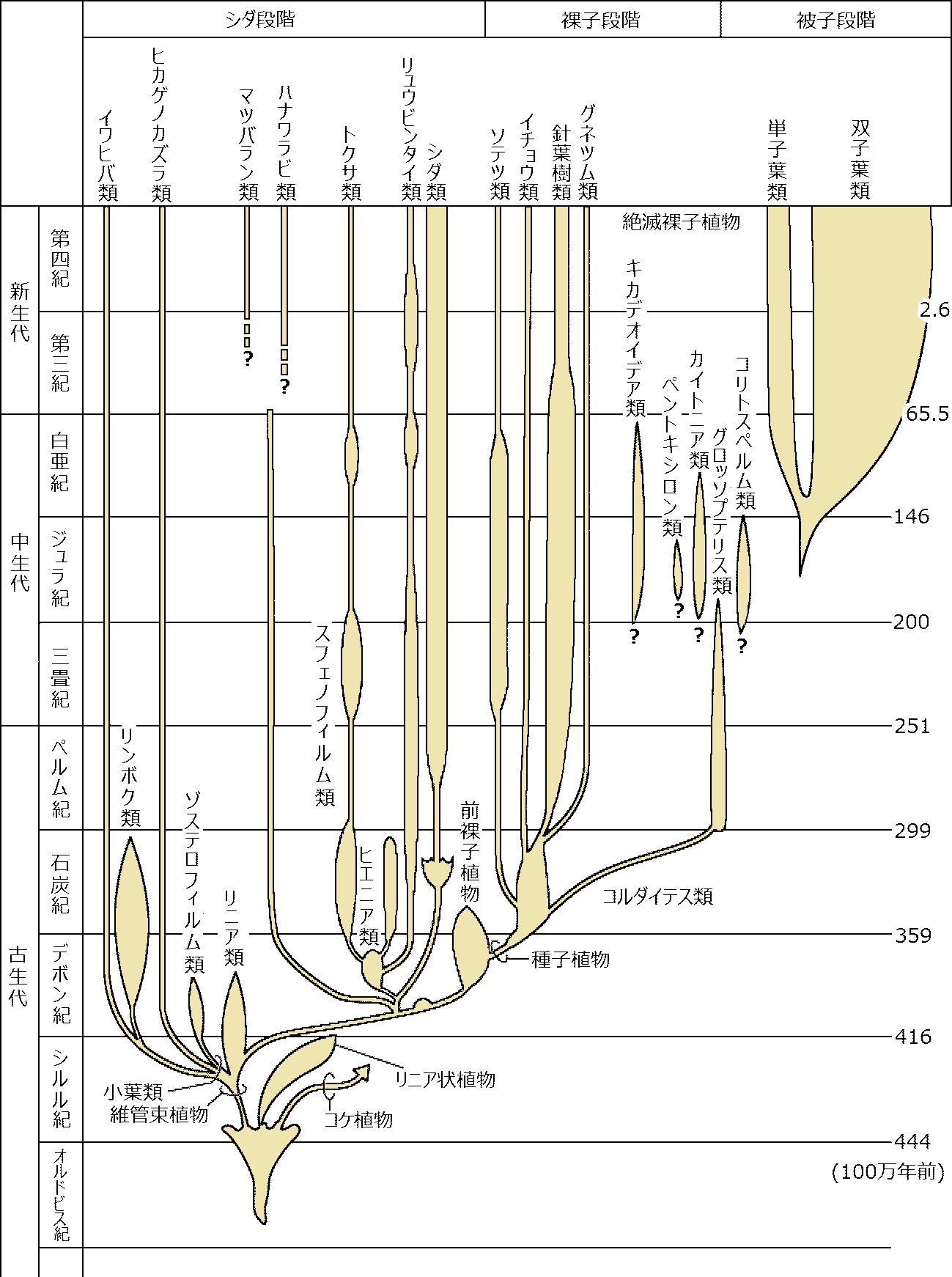 主な維管束植物の出現年代 (伊藤 2012) 巨大化石 macro-fossilカンブリア紀-オルドビス紀Aldonophyton: 陸上植物特性有するが藻類・動物の可能性もあるPridolian or Downtonian (400-395 mya)Cooksonia: 維管束植物最古の化石原始的sporocarp有 - 初期陸上植物体制は単純 primitive sporocarp → dichotomous branching (dichotomy) デボン紀 Gedinnian (395-390 mya)Cooksonia, Zosterophyllum: slit, spiral spore気孔: gurad cellがある → [進化] → Taeniocorda: lateral デボン紀シーゲニアン Siegenian (390-374 mya)Dichotomous branching → Monopodial branching (developing axis)Leaf-like organ or follicle (origin of macrophyll or microphyll) → venation, telom Development of stele and/or protostele = wood → 中心柱 stele: 外原形、中原形、内原形の3タイプ出現 Botryopteris (古生シダ): 種によって維管束形成が外原型exarchと内原型endarchをとるものがある → exarch, endarchは二次的な形質? 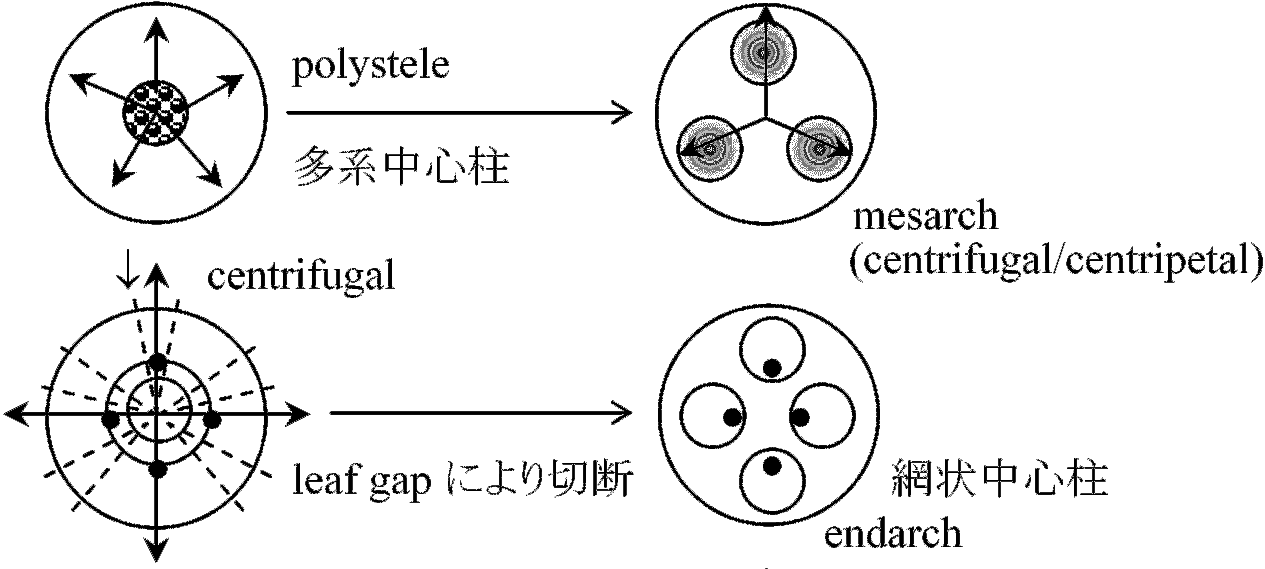 Telomeで中心柱進化は説明可 → Psilopsidaは一系統と考える
Telomeで中心柱進化は説明可 → Psilopsidaは一系統と考える
Merker (1957-58), Pant (1960): 枝につく造卵器 → Lemoigne (1967-68): Merker, Pantを支持 Sporocarp → Cone(?)デボン紀エムシアン Emsian (374-370 mya)葉平坦化し癒合起こる |
チャート岩中から化石産出: Rhynia, Psilophyton, Taeniocrada, Cooksonia, Sporogonites
体制: Cooksonia_____Rhynia________Sporogonites
1. EM観察: 横走茎(匍匐茎か葉状体)に造卵器形成 + 若胞子体胚存在
⇒ 横走茎 = 配偶体 + 直立体 = 胞子体
3. 単純維管束系存在: 単純すぎコケのcentral strandに似る。縦断面は単純な初期的螺旋構造も見られる シダ植物門 Division Pteridophyta前期デボン紀以前: 「テローム植物」は、根・葉なく、高さ1 m程度その後数千万年間: 複数群で根・葉が独立進化 + 二次肥大成長 ⇒ 巨大な体を維持可能に進化 森林誕生: 3.8 Byaまでに「森林」誕生 - 優占種は小葉植物や羊歯植物裸茎植物亜門 Subdivision Psilophytina小葉植物亜門 Subdivision Lepidophytina無舌綱 Class Aglossposida有舌綱 Class Glossopsida原シダ亜綱 Subclass Primofilicidae*デボン紀中期以降、石炭紀繁栄Angiosperm, Gymnosperm程大繁栄しない(= 地球上覆うほどではない) 概して高温好む傾向 Ex.シダ係数 pteridophyta index 一般的形態茎 stem: 短縮、根茎 – 地表面に茎殆どない(鱗片有無、形態も種の特徴)葉 leaf 単葉______羽状複葉
Order Protopteridales* (古生シダ) デボン紀-石炭紀1970 Beck CB: 殆どがCladoxylales (Cladoxylalesに移動)Protopteridaceae (古生シダ) Protopteridium (= Rellimia) Order Cladoxylales* (クラドキシロン) デボン紀中期-石炭紀胞子体のみ(×): 地上茎直立、叉状-不規則分枝、先端に樹冠形成
大部分が古生裸子植物 ≠ 古生シダ ⇔ 中心柱複雑: 分類再考必要 Pseudosporochnaceae Pseudosporochnus (Stur) Potenié et Bern. Order Coenopteridales* (コエノプテリス) デボン紀中期-二畳紀胞子体のみ: 最終分枝先端に胞子嚢特殊な原生中心柱。葉状体立体(三次元)的。葉身なし(例外あり) Aracnoxylaceae Iridopteridaceae Stauropteridaceae Stauropteris: 異型胞子 - 大胞子嚢中に2-6個の大胞子 Zygopteridaceae Botyopteridaceae: Botryopteris Renault Anachoropteridaceae Order Archaeopteridales* (アーケオプテリス) デボン紀 Devonian分類上の位置不明(暫定 有節植物)外見は現世植物的だが、葉の作りは羊歯 デボン紀に種子植物出現の証拠は今の所ない(石炭紀にはある) |
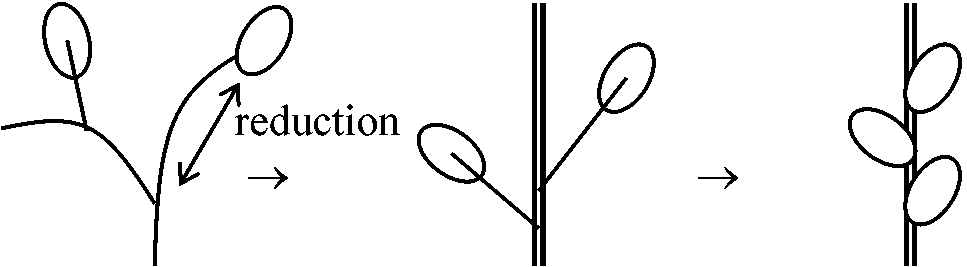
0) 葉集合説 phytonic theory葉が集合 ⇒ 茎 Ex. Tmesipteris (マツバラン): 茎成長点見られない⇔ Rhynia等原始的植物は葉がなく茎のみ → この説で説明困難 1) 単系説 (単系で陸上植物進化説明可) =テローム説 (Zimmermann 1952, 1959) + 突起説 (Bower 1935)仮定: Rhynia = 全陸上植物祖先系(単系) a) テローム説 telome theoryテローム telome: 二叉分枝した末端小枝を植物体構成単位とみなしたものRhynia: 葉がない → 大葉・小葉両方を統一的に説明しようとする デボン紀原始的葉: 発生組織注目 → "葉は軸が変化した" = 植物本体は軸 Rhynia, Hicklingia: basic morphology = 二叉分枝 dichotomous branching, sympodial types (apical cells) 二叉分枝する基本形 telome ⇒
シダ・トクサ: telomeで説明可能 ⇔ ヒカゲノカズラ: 化石上知見もなく論争中
b) 突起説 enation theory
Ex. ヒカゲノカズラ類 突起内部に維管束(葉脈)形成 + 光合成能獲得 → 小葉形成 = 小さく単純形態の葉 (小葉起源説明に適) Asteroxylon = ヒカゲノカズラ類起源? 2) 多系説シダ植物 = (無葉類 +) 有節類 + 小葉類 + 大葉類 ≠ 一系統∵ 形態的差大 → 独立祖先から陸上植物に進化 成長遅滞原則 growth retardation (Asama 1962): 成長条件↓ → 環境適応
気温年較差漸増 ☛ カタイシア植物群
植物体小型化 ⇒ セグメント(小枝・小羽片等の最小単位) |
複葉 → [癒合] → 単葉(セグメント大型化 → 枝は脈形成)
Emplectopteris: 三回羽状複葉 → Gigantonoclea: 二回 →
Bicoemplectopteris: 一回 → Tricoemplectopteris: 単葉 Alethopteris (三回羽状複葉 → 二回 → 一回) → Teniopteris 先端のセグメントのみが拡大 - 葉としては小型化 Ex. 裸子植物 = ソテツ系 + 針葉樹系: (デボン紀-)石炭紀出現茎の構造: 球果系 = 多木質, 材固い ⇔ ソテツ系 = 多髄質, 材軟らかい 1960, 62, 70, 71 Beck: 古生裸子植物
Archaeopteris (上部デボン紀標準化石): 大型羽状複葉で同型胞子か異型胞子 = シダ植物
2回羽状複葉の葉軸や羽片軸と考えた部分 → 二次木部形成 ノエゲラシア群 Noeggeranthiales古生代末期。シダ段階維管束植物化石 (系統関係未詳, 有節植物近縁?)Noeggeranthiaceae: Noeggerathia Sternb. Tingiaceae: Tingia Halle, Tingiostachya Kon'no Division Progymnospermophyta 原裸子植物門= Progymnospermopsida 原裸子植物 (> 上部デボン紀)シダ類と比べ水依存性大きく低減 系統: Cordaites (コルダボク群)につながる(?)が、まだ胞子植物段階 新赤砂岩層: 植物化石含む Order Aneurophytales (アニューロフィトン)Aneurophytaceae: Tetraxylopteris, Rellimia, Aneurophyton, Eospermatopteris
Order Pityales (ピツス)Archaeopteridaceae (Pityaceae)Archaeopitys Scott et Jeffrey (Gr. archaeo = ancient, pteris = fern): 木本 少なくとも2種は異型胞子 [大胞子 8-16 spores, 35 μm φ, 小胞子 > 300 spores, 35 μm φ] Archaeopteris Dawson (デボン紀末期)Callixylon Zalessky: Callixylon: 仮導管(階紋仮導管 saulariform tracheid)
孔紋仮導管 pitted tracheid = ナンヨウスギ ⇒ 異型胞子植物はデボン紀上部以降出現 → 現世シダは全て同型胞子 茎 = 木本, 葉 = 羊歯, 異型胞子 = 維管束植物的 Order Protopityales (プロトピチス)Protopitys Geopp.羊歯的葉 fern-like foliage羊歯植物化石分類: 分類・系統的位置付けできていない = 形態属 |
|
種子の起源: 裸子植物 gymnospermae vs 被子植物 angiospermae
単系説(単系 → 多様化) vs 多系説 (Ex. 三系説 = 大葉 + 小葉 + 有節) 同型胞子と異型胞子 homosproy and heterospory同型胞子 homosproyデボン紀: 同型胞子 50 μm φ Ex. Rhynia, Psilotum, Tmespteris (原始的)
⇒ 50 μm / 100-200 μm = 異型(二系)胞子 異型胞子 heterospory1. Licopsida: デボン紀下部Lepidostrobus: 大胞子数減少、大きさ増す Lepidocarpon (石炭紀): 胞子嚢に1個の大胞子 ≈ 種子 Lepidodendron: 器官に包まれる = megasporangium 2. Spheropsida: デボン紀上部 - Calamostachys3. Progimnospermae: デボン紀上部 - 針葉様木本 4. 胞子散布: シリル紀上部 ⇒ 種子の起源は多系? (同型胞子 → 異型胞子 → 種子) シダ状種子植物 seed ferns1903 Oliver F & Scott D: シダ状種子植物(Pteridosperms)提案Archaeosperma: デボン紀末、最古種子様化石(他石炭紀以降) = 石果 幹: Lyginopteridales(真正中心柱型)/Medullosales(多条中心柱型) Lyginopteris: 樹状シダ種子の茎 = Sphenopteris 葉 + Lagenostoma ♀繁殖器官 + Crossotheca ♂ 1963 Archangelsky: パタゴニアの白亜紀前期の植物化石
多くのシダ状植物化石はシダ種子植物 |
1968 Pettit & Beck: 種皮 seed coat確認
種子は不完全な盃状体に囲まれ、種子先端は分裂し4個の大胞子はシダ植物の様に四面体状に配列
種子 seed癒合 seeds in fruit: integument → seed coat起源
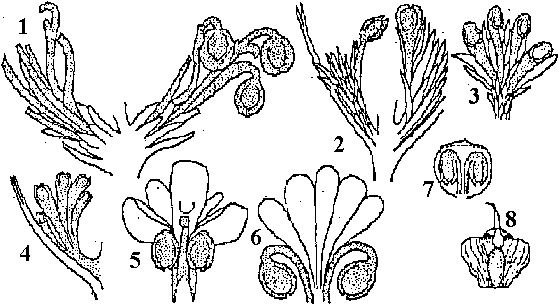 Fig. Cordaianthus → 針葉樹 (胚珠構造単徐々に純化)。1. Cordaianthus, 2. Lebachia, 3. Wallchiostrobus, 4. Ernestiodendron, 5. Pseudovoltzia, 6. Cryptolepis, 7 Hirmeriella, 8. Araucaria
Fig. Cordaianthus → 針葉樹 (胚珠構造単徐々に純化)。1. Cordaianthus, 2. Lebachia, 3. Wallchiostrobus, 4. Ernestiodendron, 5. Pseudovoltzia, 6. Cryptolepis, 7 Hirmeriella, 8. Araucaria小葉系: Protolepidodendron (デボン紀) → Lepidodendron (石炭紀) →
Lebachia (石炭紀後期-二畳紀): 雌雄別胞子嚢穂 (直立 = ♀ 横走 = ♂) Podocarpus (現世) |
☛ 地質年代 中生代
三畳紀 Triassic 2.51-1.95 Bya中生代ユーラシアフロラ: 植物群組成貧弱Ex. シダ類、ソテツ葉類、Voltzia(球果綱), Schizoneura(トクサ綱) Pleuromeia: ヒカゲノカズラ綱 三畳紀初期ユーラシアフロラ表徴的種
茎分枝しない。2 m h, 9 cm φ ≈ レートライアス型植物群(三畳紀中期未発見の頃の呼称) ヤブレガサウラボシ Dipteridaceae: 多種多様(現生1属Dipteris8種) 葉身に網状脈: Thaumatopteris, Dictyophyllum, Clathropteris, Camptopteris, Hausmannia, Goeppertella 汎世界的分布 (↔ 三畳紀初期: 分布地域限定的) ジュラ紀 Jurassic 1.95-1.35 Bya日本1961 浅間: 後期ジュラ紀-前期白亜紀に植物区系手取植物群 Tetori-type flora: 上部ジュラ系-下部白亜系, 1.7-0.96 Bya = 内帯植物地理区 ≈ シベリア植物地理区
シダ(タカワラビ科),イチョウ、広葉球果 下位 = 九頭竜: ジュラ紀後期、分布狭い 上位 = 石徹白: 下部白亜紀系部分 = 尾口層 + 赤岩層 尾口層: 化石多産 Ex. 白峰村桑島化石壁
沼沢地 → 流入河川沿に汽水産、淡水産貝類化石
中生代古期型植物の系統化石多産 (領石植物群: *未報告 §稀) Coniopteris, Birisia, Dicksonia, Eboracia
ソテツ綱(古期型): Ctenis優勢 + Nilssonia* Ginkgoidium, Eretmophyllum, Sphenobaiera, Pseudotorellia 球果綱: Podozamites*優勢§: P. reinii, Xenoxylon, Pseudolarix, Sequoia, Parataxodium, Athrotaxites, Athrotaxopsis, Cephalotaxopsis トクサ小型化 ≈ 現生種形態 領石植物群 Ryoseki-type flora: 上部ジュラ系-下部白亜系, 1.6-0.96 Bya= 外帯植物地理区 (ウィルデン型)
東北-九州の太平洋側 (1890 Nathorst AG) (手取植物群: *未報告 §稀)
+ Gleicheniaceae広分布 ソテツ葉類: 普通 Zamites§ (Zamiophyllum buchianum), Ptilophyllum§ Nilssonia - 小型で細長い葉§ |
イチョウ葉類: 極稀 後期ジュラ紀-前期白亜紀1964 Vakhrameev VA: 北半球フロラに2型 = 古期型 + 新期型
古期型 炭層発達、温暖・適湿
| ジュラ紀中期: 古期型・新期型混合 Ex. 舞根植物群| 白亜紀 Cretaceous 1.4-0.65 Byaイネ科: 化石資料少
白亜紀後期恐竜糞化石中 - プラントオパール(イネ科特有)発見 被子植物 angiosperms 起源 origin被子植物: 白亜紀初頭(140 Mya)に化石からは突然出現ポルトガル、バージニア-カナダ西部、シベリア北東部-沿海州 1932 Harris: Furcula ⊂ 被子植物 (否定: ⊂ シダ種子)
グリーンランド上部三畳系 コロラド州サンミゲル渓谷上部三畳系 1975 クラシロフ: 古生被子植物綱提唱Dirhopalostachys = Nilssonia schmidtii (Heer) Seward. ♀ 白亜紀末: 形態・分布 ≈ 現在
裸子植物より世代交代早い、環境順応性高、動物と共進化 北極起源説 [否定] 植物相: 東亜 ≈ 北米東海岸 → 北極起源で説明可 1940 Vavilov: 遺伝子中心説 = 作物起源1966 Takhtajan AL: 東南アジア熱帯域起源説 - 地域の特徴 原始的植物多 + 植物相多様 + 環境複雑 1960 前川: 古赤道分布説 [否定]1970 Axelrod: 赤道圏山地発生説 - 多様な環境下で突然変異率は高い 1976 Kreutzer: 南極起源説 [否定] 南極 - [移動] - パタゴニア・オーストラリア・南アフリカ 白亜紀後期: 被子植物爆発的分化・繁栄 - 世界広分布日本 足羽植物群 (松尾 1962): 上部白亜系(手取植物群上位) - 半数現世類似種 函淵植物群(ニルソニア層): 白亜紀末 - Nilssonia豊富
1910 Stopes & 藤井: 被子植物6種他 |
|
化石推定殆ど: 中生代地層は断片的だが世界各地に見られる この時代には地球全体のフロラに相違が余りない 気候: 温暖安定(古生代後期、特に石炭紀以降) → Alps orogene, circum-Pacific orogene: 中生代(特にジュラ紀) [内陸] 湿潤 → 乾燥 → (Variscan orogeny) → 温和_________→ Gymnospermae, Ferns繁栄 多種が気候激変のため白亜紀下部-上部にかけて絶滅 逆に多くの被子植物現れる(1億2500万年前) - 動物との共進化重要
Cretaceousに確認され、現在まで続くグループ
湖畔: シダ類・裸子植物 中生代種子植物Division Spermatophyta 種子植物Subdivision Gymnospermae 裸子植物Class Cycadopsida ソテツSubclass Pteridospermidae* シダ状種子植物特徴: 羊歯状葉状体に裸子種子形成Order Pteridospermales シダ状裸子植物 Order Bennettitales*: 中生代三畳紀-終期 Order Cycadales ソテツ Nilssoniaceae*: ジュラ紀-白亜紀 Ex. Nilssonia Subclass Pentoxylidae ペントキシロン*Order Pentoxylales*Class Coniferopsida 球果植物Order Cordaitales: 上部デボン紀-二畳紀。北半球多出1931 Elias: Equisetumで種子報告 ムカゴと訂正 (現世も時々ムカゴつけるEquisetum発見される) 1957 Boxter: Carpannularia = イワヒバ型発生をする胞子嚢 sporangiumしかし、integument形成しない ≠ 種子 EristophytaceaeCordaitaceae* 石炭紀-二畳紀 Amyelon Williamson (根), Cordaianthus Grand'Eury (球果), Cordaicarpus Geinitz (種子), Cordaites Unger (葉), Mesoxyllon Scott et Maslen (茎) Cordaites = 球果(石炭紀 3.5 Bya) ⇔ イチョウ・ソテツ(3(2.8) Bya)
球果: 雌雄別。葉: 樹冠部枝に螺旋状配置、平行脈 Order Coniferales (Pinales) ジュラ紀 Jurassic: 化石豊富(日本はジュラ紀化石産出しない) - 南半球フロラは比較的均一。裸子植物繁栄 Angiospermae 被子植物: 白亜紀Cretaceous初期以降出現(化石産出) → 昆虫出現と密接に関連
裸子植物 → [乾燥適応] → 被子植物 1. 単純葉Ginkgo, Conifer, etc.(現代)a) ramification (branching)
venation 前2種がGinkgoの起源、Czekanowskiaのみが雌器官を有しており、前2種に非依存的である b) linear-leaved plant with parallel venation
Elatides, etc. |
2. 胚珠 ovule ☛ エングラー体系閉じていないcarpelに囲まれるMagnoliaceae, Cercidiphyllaceae, Nymphaeaceae – 隔離分布 表. Magnoliales(モクレン)及び近縁原始的被子植物の分布型 - 環太平洋を取り巻くよう分布 [北半球型] Magnoliaceae (モクレン)Trochodendraceae (ヤマグルマ): 東アジア(日本-台湾、朝鮮南部)、固有 Tetracentraceae (スイセイジュ): 東アジア(中国大陸南部-東ヒマラヤ)、固有 シキミ Illiciaceae: 東アジア-マレーシア西部(高地)、北米東部-中米高地、キューバ マツブサ Schisandraceae: 東アジア-マレーシア西部、北米東部 [熱帯型] センリョウ Chloranthaceae: 中南米、東アジア-マレーシア-太平洋南西部バンレイシ Annonaceae: 汎熱帯、北米東部 ニクズク Myristicaceae: 汎熱帯 クスノキ Lauraceae: 汎熱帯-亜熱帯、北半球では温暖帯まで カネラ Canellaceae: 熱帯アメリカ、アフリカ、マダガスカル [南半球型] シキミモドキ Winteraceae: マレーシア、豪州東部、NZ、南米-中米、マダガスカルデゲネリア Degeneriaceae: フィジー(南西太平洋) ヒマンタンドラ Himantandraceae: 東豪州、ニューギニア、モルッカ エウポマチア Eupomatiaceae: 東豪州、ニューギニア アウストロバイレヤ Austrobaileyaceae: 豪州(北東部) モニミア Monimiaceae: 南米、アフリカ-マダガスカル、マレーシア-豪州 3. 花 flower単純One stamen – One ovule Amentiferae: pollen = tricolpate - primitive (Engler 1879) Magnoliaceae: monosulcate pollen ⇒ モクレン科が被子植物の起源(田村 1974) 4. 葉脈 veinCycadophyte(ソテツ状葉印象化石類)平行脈 – 被子植物起源?(Bennetiales) Ginkgophyte: 叉状脈Coniferophyta 被子植物は様々な所に雑草的に出現 → 徐々に広がる 被子植物の特徴
花粉 pollen被子植物花粉化石 + 白亜紀中期出現(多様性低 - 以降多様な花粉産出)表面網目模様 areolate ⇒ 1) 虫媒 2) 花粉が柱頭に付着しやすい という2つの利点を得た進化 受粉: 受粉完了し柱頭着床検知するにはmonosulcateよりもtricoplate, tricolporateの方が優れるMonosulcate: 溝が横を向くと着床を検知しても花粉管が到達しにくい 花粉孔の形態 縦/横 > 2 = 溝型
長口型 sulcate type 溝型 calpate type 孔型 porate type
網目模様のない花粉出現: 風媒、花粉が小さくなる → 当時の気候が一時的に乾燥? – 虫媒効率低下
|
|
古生代: 三葉虫 trilobite・サンゴ coral・腕足類 brachiopod 三葉虫 Class Trilobita (Fortey 1997): カンブリア紀-ペルム紀(二畳紀)末 Order Agnostida; カンブリア紀-オルドビス紀 Order Redlichiida: カンブリア紀 Order Corynexochida: カンブリア紀前期-デボン紀中期 Order Asaphida: オルドビス紀-デボン紀中期 Order Phacopida: カンブリア紀末-デボン紀後期 Order Proetida: オルドビス紀-ペルム紀末 Order Ptychopariida: カンブリア紀-デボン紀後期 Order Lichida: カンブリア紀後期-デボン紀中期 ウミユリ (棘皮動物), sea lily 最古化石: バージェス頁岩中(カンブリア爆発時出現)、破片化石多、浅海底 = 銭石(赤坂金生山)、千貫石(東北)、梅花石(九州) シルル紀: 5 bya 無顎類(≈ 甲冑魚類)出現 → 4.4-4 bya 化石多産鎧状の甲羅 - ウミサソリ(Eurypterus)に対する防御? シルル紀後期: 有顎類出現 → デボン紀発展放散(板皮類 - デボン紀末絶滅)デボン紀: 総鰭類 - 上陸 → 古生代末期衰退 シーラカンス (総鰭類) - 生きた化石 デボン紀末期: 両生類 - 古生代後期繁栄デボン紀化石: 側線器官 - 魚類的 Class Reptilia 爬虫綱 (爬虫類 reptiles)単弓類 Synapsida: 哺乳類起源Superorder Dinosauria 恐竜 (s.s.), dinosaur古生代(石炭紀), 3 bya - 双弓類(≠ 恐竜)起源中生代: 爬虫類時代 三畳紀 (P/T境界後): 恐竜出現 → 爬虫類繁栄 - 乾燥(砂漠時代) ジュラ紀: 巨大化 白亜紀: 最盛期 [被子植物出現] → 終期: 衰退 → 昆虫、小型動物に代わる 1923 ゴビ砂漠で恐竜卵化石 → Protoceratops 以下2目(竜盤・鳥盤)が分類学上の恐竜Order Saurischia 竜盤Suborder Theropoda 獣脚類二足歩行 (≠ ヒト二足歩行) + 前肢退化
草食恐竜 + 大型化 Saturnalia: 中生代三畳紀後期、南米 Thecodontosaurus: 三畳紀後期、グレートブリテン島 Infraorder Prosauropoda 原竜脚類 Anchisauridae: Anchisaurus Plateosauridae: Plateosaurus Melanorosauridae: Melanorosaurus Infraorder Sauropoda 竜脚類 Omeisauridae: Mamenchisaurus Dicraeosauridae Diplodocidae: Apatosaurus, Diplodocus Camarasauridae: Camarasaurus Brachiosauridae: Brachiosaurus ブラキオサウルス Titanosauridae: Titanosaurus), Saltasaurus Order Ornithischia 鳥盤恒温動物説: 現世爬虫類 = 変温動物 → 動き遅い骨格構造 → 恒温動物(全恐竜にあてはまるわけではない) Anatotitan Chapman et Brett-Surman 1990, syn. Trachodon (廃棄)Nipponosaurus Nagao 1936 1924 ニッポン竜: 樺太、白亜紀後期 1962 長崎高島炭鉱地下917 m、当初カモノハシ竜 → 哺乳類 Archosauria (恐竜群/主竜群, s.l. 絶滅)心臓2心房2心室Order Ichthyosauria 魚竜2億5千万年前出現-9000万年前絶滅Elasmosauridae Futabasaurus suzukii Sato, Hasegawa et Manabe 2006 1968 化石発見 (フタバスズキリュウ/双葉鈴木竜) - 首長竜日本初確認 福島県双葉層群玉山層入間沢部層河岸露頭 2006 新属新種として記載Order Protosauria 原竜Order Mesosauria 中竜, mesosaur (middle lizards)古生代後半。この分布をもとにゴンドワナ大陸復元Order Paraspida三畳紀初期から出現(Super)Order Sauropterygia 鰭竜Order Nothosauria 偽竜, nothosaur三畳紀 (→ 首長竜に分岐)
Nothosaurus Hunter 1834 1939 宮城県本吉郡柳津町北上川岸中生代三畳紀地層から化石
体の大部分保存 (頭骨、四肢骨不明) - 戦後紛失。全長 ≈ 1.3 m Order Eosuchia 始鰐Order Thecodontia 槽歯恐竜群で最も原始的 - 原始的が共通なだけで有効分類群ではないProtosuchus, Telosaurus, Alligatosaurus ⇒ 爬虫類の起源? Order Pterosauria 翼竜, wing lizard1784 Cosimo Alessandro Collini (1727-1806, 伊): 化石初記載Rhamphorhynchus: ジュラ紀 Pterodactylus: ジュラ紀後期 Pteranodon 翼手竜(プテラノドン): 白亜紀、北米 Quetzalcoatlus: 白亜紀、北米 Order Therapsida 獣形Order Ictidosauria イクチドザウルス爬虫類-哺乳類中間形態節足動物鳥類1861 始祖鳥(ドイツで発見): 三畳紀, 爬虫類-鳥類中間
鳥類特徴: 前足が翼。体表に羽 |
哺乳類有袋類(哺乳類): ジュラ紀ウマ (horse): 化石連続的変化 (ウマの進化 ♠: 草食, ♥: 木の実食)
× Equus 150, 1/1 ♠ 最新世 ×
└───┬────┘ ┌────┘
(*: 絶滅) ☛ ゾウ(現存) Elephantidae (ゾウ, elephant)瀬戸内海: 象化石の宝庫Palaeoloxodon (ナウマンゾウ): P. naumanni, Naumann's elephant
誕生: アフリカ(鮮新世) → 拡大: ユーラシア(更新世) Mastodon マストドン* Stegodon ステゴドン* S. aurorae (アケボノゾウ), syn. S. akashiensis (アカシゾウ) ⊃ S. kwantoensis (カントウゾウ), S. sugiyamai (スギヤマゾウ), S. akashiensis (アカシゾウ), S. infrequens S. orientalis (トウヨウゾウ)Loxodonta africana africana アフリカゾウ Elephas maximus アジアゾウ(インドゾウ) Mammuthus Brookes 1828 マンモス* M. africanavus アフリカ-, M. armeniacus アルメニア-, M. columbi コロンビア-, M. exilis コビト-, M. hayi, M. jeffersonii ジェファーソン-, M. lamarmorae, M. meridionalis メリジオナリスゾウ, M. primigenius ケナガ-, M. protomammonteus ムカシ-, M. subplanifrons, M. sungari ショウカコウ-, M. trogontherii ステップ- 北広島マンモス: ケナガマンモスとナウマンゾウの臼歯発見 = (可能性) 4.5万年前共存 + 針葉樹林帯生息 (≠ 定説) (北広島マンモス大復活プロジェクト, 北海道博物館・北広島市エコミュージアムセンター企画) ビラフランキアン期 Villafranchian age: イタリア北部陸成層に示される時期
ゾウ,ウマ,レイヨウ出現
mastodon 第三紀型ゾウ、他 第四紀型ゾウ Order Desmostyliformes 束柱デスモスチルス。海牛類や長鼻類と近縁(desmos束 + stylos柱) (! = 絶滅)中生世前-中期の北太平洋沿岸のみ 切歯円束形。臼歯高歯冠柱状歯。大臼歯水平交換。側頭骨頬骨突起が上顎骨と接触 パレオパラドキシアから分布北に偏り寒流系種。日本: 岐阜・島根以北産出 Order Desmostylida中新世!Order Condylartha 顆節原始的有蹄動物 !Order Notoungulata 南蹄原始的有蹄動物 !。南米Suborder Notioprogonia 南祖Suborder Hegetotheria ヘゲトテリウムSuborder Typotheria ティポテリウムSuborder Toxodontia トクソドンOrder Litopterna 滑距原始的有蹄動物 !。南米Macraucheniidae マクラウケニア Proterotheriidae プロテロテリウム Order Pantodonta 汎歯大型有蹄動物 !。バクに似た草食動物Order Dinocerata 恐角ウインタテリウム類。巨大有蹄動物 !Order Pyrotheria 火獣巨大動物 !。南米Order Xenungulata 異蹄大型有蹄動物 !。南米Order Embrithopoda 重脚アルシノイテリウム類。大型有蹄動物 !。エジプト地方霊長類 primates第三紀 = 霊長類時代暁新世中期: ツパイ 北米大陸に多くの化石 - いずれも極めて原始的
食虫類(ツパイ) → 樹上生活 → 霊長類 始新世: 現世哺乳類種の半数の化石 後期: 類人猿(様)化石 (Amphipithecus mogaungensis) - 原始的狭鼻類 洪積世前期-中期における日本の哺乳類動物群の由来(大陸と陸続き)泥川湾動物群 柳城動物群 万県動物群 周口店動物群 第四紀 = 人類の時代 |
☛ 地質年代: 新生代
第三紀 (Tertiary64.3-2.6 Mya大陸移動に伴うフロラの地域差起こる →北半球中部以北では熱帯(亜熱帯)性植物減少。落葉広葉樹が主構成要素 (現在) 共通種 北半球三大陸間: ブナ、ナラ、シナノキ、カエデ 北米-東亜: ユリノキ、スズカケノキ、カヤ、イヌガヤ、ツガザクラ 東亜: メタセコイア、イチョウ 暁新世 Palaeocene 65.5-55.8 Mya始新世 Eocene 55.0-38.0 Mya初期-中期: 白亜紀後期植物群の延長だが中生代的要素なし
高島炭田: 亜熱帯的組成 Ex. 熱帯性シダ、シュロ 夕張炭田: 亜熱帯的種見られる Ex. シュロ - 亜熱帯林・温帯林混交 北海道: ノグルミ、ハンノキ、サワシバ、ハシバミ、クリ、プラネラ、ケヤキ、カツラ、カエデ、ウリノキ、カシ、ビロウ、バショウ、メタセコイヤ、スイショウ、ラクウショウ、イヌガヤ、マツ(Abies, Picea, Tsuga 亜高山地帯の花粉) [常緑広葉樹主体。若干針葉樹。現在の南九州以南林相に似る]
極地、グリーンランド、スピッツベルゲン: 亜熱帯・暖温帯植物 漸新世 Oligocene 33.7-23.8 Mya北海道: クルミ科、カンバ科、ニレ科、カツラ科、マンサク科、カエデ科、バショウ、コモチシダ、イヌワラビ、コウヤワラビ、ゼンマイ、メタセコイヤ、スイショウ、ラクウショウ [落葉広葉樹主としメタセコイヤ交える針広混交林]中新世前期まで亜熱帯林南下 ⇒ 種入替り現代的種増える 新第三紀 Neogene (Neo-tertiary) 25-2 Mya北半球第三紀植物相の遷移: 種レベルで分類可 + 分布は現在とかなり近い要素: 1 ヨーロッパ, 2 コルシカ, 3 東亜, 4 北米東部, 5 北米西部, 6 寒帯 寒冷化傾向 (第四紀更新世まで) ⇒
→ 植物移動は砂漠、海、山脈等で遮られる 中新世 Miocene 23-5 Mya北海道[前期] ☛ 中新世 温帯・冷温帯落葉広葉樹主体 + 球果類(高山) ↔ 常緑広葉樹少 現日本自生種より中国中部や北米東部自生種の近縁種多 Salix, ポプラ, サワグルミ, クルミ, ハンノキ, カンバ, クマシデ, ハシバミ, アサダ, ブナ, ニレ, ケヤキ, カツラ, トチノキ, ナナカマド, サンザシ, カエデ, ウリノキ, メタセコイヤ, スイショウ, モミ, ハリモミ, ツガ, アスナロ属 [中期] (20 Mya) やや温暖 → 落葉広葉樹林 ノグルミ, サワグルミ, ハンノキ, クマシデ, ナラ, クリ, ケヤキ, シンジュノキ(× シンジュ), Acer, ウバメガシ, ツバキ, ナツメ, メタセコイヤ, スイショウ, コウヨザン, タイワンスギ, イヌカラマツ, クロマツ, モミ, ハリモミ, ツガ, トガサワラ属 [後期] (15 Mya) メタセコイヤ絶滅。落葉広葉樹林と共に針葉樹増加 ≈ 現在の森林 Salix, ドロノキ, クルミ, サワグルミ, ミヤマハンノキ, ダケカンバ, ウダイカンバ, シラカンバ, ブナ, カツラ, ニレ, イタヤカエデ, ハリギリ, シナノキ, マメ科, バラ科, ホウノキ科, フウ, シンジュノキ, ナツメ, タイワンスギ, メタセコイヤ, Picea, Abies (A. firmaとは同定できない), Tsuga |
鮮新世 Pliocene 5-2.58(1.6) Mya1967 吉山: 北浅川(八王子)河床でメタセコイア化石林発見 - 鮮新世後期林床は泥炭層 Metasequoia: 白亜紀-新第三紀繁栄-第四紀氷河期衰退1941 三木: Sequoiaから独立 - Metasequoia命名 当初同定: 葉 = Taxodium, 果実 = Sequoia → 同一種 1944 Wang: 生きたメタセコイア発見 - 生きた化石 (詳細研究可能となる)中国四川省磨刀渓上流祠近く。幹2.3 m φ - 各地に播種・移植 第四紀 Quaternary 2.588 Mya-現在初期 early Quaternary = 氷河期 → 赤道付近に植物集中各大陸で個別に種分化 更新世 (Pleistocene) 2600 Tya (2.6 Mya)-11.5 Tya
山脈方向 氷期-間氷期植物移動 最寒冷期 → 垂直分布は現在より1000-1500 m低 (針広混交林だが、寒暖の反復に対応しつつ現在の森林相に近づく) 完新世 (Holocene) 11.5 Tya-現在欧州: ツンドラは消滅、ステップは森林化(北極域にも森林発達)
|
|
(現在含め)地球史や気候条件変遷 → 生物進化・移動 → 地史学的探求
Ex. 花粉分析、珪藻、sedaDNA 珪藻珪酸質殻 = 堆積物中残りやすく化石から湖・湿原化学環境(一次生産)推定一般的手順
堆積物古代DNA (sedaDNA)過去の生物の遺伝物質が堆積物中に保存されたもの ⇒
化石・花粉分析で発見困難な微細生物痕跡検出可 → 生態系・気候変動 A. 第三紀以前気候的環境変化を考える際の基礎概念
1. 大陸移動 continental drift 2. 極移動 polar migration B. 更新世の環境地磁気と絶対年代の対比による時代考証植生変遷: 花粉分析 pollen analysis 後期: 北海道 – マンモス。東北地方花泉(=花泉階) – ナウマンゾウ、ヤギュウ等。新潟 – トナカイ
暖帯要素出現 中期: ナウマンゾウ、ニッポンムカシジカ、楊子シカ、マチカネワニ 出現、滅亡の時期は断定しにくく、繁栄、衰退の判断も難しい。例えば、種類数で判断するのか、個体数で判断するのかなどという化石化の問題がある。始祖鳥 Archaeopteryx の化石は5片しか見つかっていないパレオソル(古土壌)2r + 2H2O > 2rO2 + H2 |
古生物地理区 paleobiogeogrpahy生態群集を遺骸群集から復元
↓ (生物)群集 Biocoenosis 生活型 habitat type生活型によって古環境を推定復元する (functional morphology)
化石の配列dimensionの方向等の化石形成因子を考慮する必要がある 地質学(geology)との関連 (relationships to geology)珊瑚礁 – 遷移段階と対応(地質学的遷移及び生態学的遷移)植物群集、動物群集が基盤(地形)形成 → その上に群集発達 + 垂直分布にも注目すべき 石灰岩 sparite
バイオスパーライト biosparite: 生物の堆積 spar 古赤道 paleoequator古い時代(特に白亜紀から第三紀)の赤道の位置 - 地軸が現代と異なる古赤道に沿い分布: ドクウツギ (Coriaria japonica)・スミレ・アケビ科
|
|
多くの湖底・泥炭堆積物: 花粉・胞子遺体含む 他遺体・化学成分・同位体測定結果等とあわせ古環境復元計る 花粉分析 pollen analysis(堆積物中の)花粉の種類と量を調べること花粉胞子分析 pollen-spore analysis: 羊歯類・蘚苔類胞子も調べた場合 層順が明らかなことが不可欠(+ 絶対年代)。湖底堆積物を5 mm間隔で調べれば遷移解析可能
花粉 (1)模様(表面・断面)、(2)孔(発芽孔)、(3)構成物質、 (4)花粉管、等の特性利用 野外採取コンタミ避ける Ex. 採取用SS型ピンセットは頻繁にクリーニング原理と方法花粉・胞子は10% KOH溶液、48%HF錯化液acetolysis solutionに無反応→ 目的物以外の基質を化学的除去(HF処理 - 花粉10%小さくなる)
|
(15)から(18)の操作を錯化処理acetolysis treatmentという 日本第四紀堆積物調査で知るべき花粉: モミ、トウヒ、ツガ、マツ、カラマツ、ビャクシン、ナギ、コウヤマキ、スギ、イチイ、ブナ、カシ、ナラ、カンバ、ハンノキ、ハシバミ、シデ、ヤマモモ、ヤナギ、ケヤキ、ニレ、シナノキ、クルミ、サワグルミ、ヨモギ、ブタクサ、カラマツソウ、オオバコ、ワラビ、ヤマドリゼンマイ、ヒカゲノカズラ、イワヒバ各属とイネ、スゲ、キク、アカザ、キツネノボタン各科 現植生花粉分析による情報表面試料: 湖底堆積物や腐食土極表層花粉集団 = 過去10数年堆積物 → 過去花粉構成解釈時の基礎資料温暖帯: 照葉樹林 laurelled forest, Lauriginosae に特徴づけられる
クスノキ科花粉: 外膜発達しない = 分解し易い - 普通遺体とし残らない 冷温帯: ブナ帯 - ブナ、ミズナラ、シラカンバ花粉 亜高山帯: ハイマツ花粉 |
= 大量絶滅 (great extinction, or mass extinction) - 少なくとも過去5回
1) オルドビス紀末 (O/S境界)4.431-4.429億年前 - 雲南省永善県におけるO/S地層境界面分析2) デボン紀末 (F/F境界)3.74億年前寒冷化 + 海洋無酸素事変 3) ペルム紀末 (P/T境界)Permian (古生代ペルム紀) - Triassic (中生代三畳紀) 境界2.51億年前 → 地球上の90%以上の生物絶滅
[地球史上最大の絶滅] 三葉虫・筆石・紡錘虫(フズリナ) 4) 三畳紀末火山活動説。隕石説5) 白亜期末 (K/T境界, C/T境界)Cretaceous-Tertiary boundary (K-T boundary)6500万年前(白亜期末) → 約50%が絶滅
恐竜 + アンモナイトや海洋原生生物 → 絶滅
アルヴァレツ(地質学者) → K/T境界発見 |
イタリア以外にIrを多く含むK/T境界層見つかる + クレーター発見 絶滅を逃れた恐竜類もいただろう 緩やかな気候変化説 = 長期絶滅
海洋化石: 原生動物やアンモナイトの絶滅は、短期間にほぼ同時に起こった → 隕石衝突説支持
火山噴火 → 冷蔵庫効果 → 陸上環境徐々に変化
プレート運動 → 大陸配置変化 → 海流変化 → 海環境徐々に変化
両説でも説明できない点もある ☛ 絶滅危惧種 6) そして現代現代の絶滅速度は、化石記録の1000倍の速さ将来の絶滅速度は、現在の予測速度の10倍の速さ → 化石記録の10000倍 |
 a
a b
b

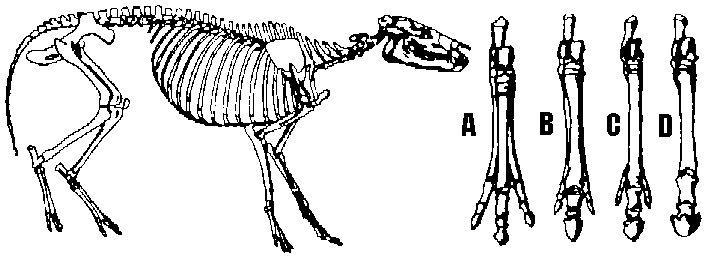 Hyracotherium (Cope ED). 肩高30 cm。背丸く首短い。指前4本後3本。奇蹄類共通祖先とも思われる。A-D: 全て右後足(大きさは同じに描く)。A: Hyracotherium, B: Miohippus, C: Merychippus, D: Equus
Hyracotherium (Cope ED). 肩高30 cm。背丸く首短い。指前4本後3本。奇蹄類共通祖先とも思われる。A-D: 全て右後足(大きさは同じに描く)。A: Hyracotherium, B: Miohippus, C: Merychippus, D: Equus