(2026年3月20日更新) [ 日本語 | English ]
HOME > 講義・実習・演習一覧 / 研究概要 > 小辞典 > 哺乳類 > 人類
|
人類学 anthropology: 人類(霊長類ヒト科)に関して研究する学問
人類学系統文化人類学 cultural anthropology (社会人類学, s.l.): 文化・風習・習慣・慣習・風俗・宗教等分化 → 原因
社会人類学 social anthropology (s.s.)
認識人類学 cognitive anthropology: 社会の事物事象分類体系化
民俗(人類)学 folklore: 文化模様
化石人類学(古人類学)
生態人類学: 人間生活に着目し、環境への適応と進化を研究 - ヒト個体群生態学という側面強い |
生態人類学 ecological anthropologyヒト anthropoid, Homo sapiens sapiensヒト = 大和言葉人間 = 仏教用語 人類 = 生物学的呼称 Ration: ヒトとは「2本足で立って歩いて、羽の無い動物」 人間脳神経系発達による行動パターン、技術体系、観念・価値体系、社会組織、情報システムが重要文化生態学中核文化: 環境適応に基本的な生業活動や経済機構に密接に関連する一群の特性食物エネルギー獲得・生産技術、人口変動、土地・資源所有・利用システム等 2次的文化: それ以外の文化特性人類社会の多様な資源利用システムとそれに関連する社会システムを進化視点から理解 地球規模での環境問題深刻化Ex. 環境汚染 (environmental pollution)Ex. 歴史: 狩猟採集民がマンモス、ドードー、モア等の大型鳥獣類を絶滅 ↔ 人類学が主要研究対象とした狩猟採集民、農耕民、牧畜民が培ってきた環境と調和した生存システムの理解が、開発と共に進行する環境破壊や資源の過剰利用の解決にヒントを与えると期待 Ex. コモンズ(エココモンズ): 土地・資源共同所有・利用、あるいは環境の不規則性や劣化に対する地域社会レベルでのリスク回避のシステム等が、多くの社会に存在(大塚 1994) ⇒ 活用し開発と環境保全の両立を模索 |
|
人間とは: 生物学的分野・社会学的分野で人間(ヒト)概念異なるのが現状 1923 臨界点学説 critical point theory (Kroeber AL) = 人の他の動物との大きな違いは大脳皮質の発達 変化は進化が突然ジャンプするよう大きく変化した(大部分否定)
= 二足歩行 + 石器使用 + (脳量 ≤ 1/2·現代人)
多くの発掘化石: 解剖学的にヒトとそうでないものの区別困難 - ヒトは徐々に進化 = 「臨界点学説」否定 → 単に自然環境による淘汰の他に文化を人間は持った事が人類進歩の点で大きな意味がある 文化は人類固有のものか日本猿: 個性存在 → (以前)チンパンジー = 最も人間に近い1) 系統図(幸島のサル)完成 → 京大餌付実験 最初はイモをそのまま食べた → あるサルが泉でイモ洗いし食べる 泉が干上がった時に、仲間(大分イモ洗いは伝播済)達は海へ降り塩水でイモ洗いを始め、塩加減が良かったらしく一口かじる毎にイモを塩水につけ食べる事を始めた → 文化? (模倣伝播) – 伝播遅い |
観察継続: 衝撃的事件発生 → 餌(麦)を砂場に撒くと(麦粒のみを選んで)砂と一所に食べた → イモを塩水につけ始めたサルが麦と砂を持ち海へ入り麦を塩水に撒き付け、砂は沈み麦は浮き選び食べた。ある日、この麦を塩水に漬けていた所、麦略奪者が表れた → 麦搾取階級発生 = 階級社会発生 京大研究: サル ≈ 30の音声信号(≠ 言葉) - 複雑多岐使用不可 ≠ 言語 2) 米国の研究 a) ヘイズ(Keith & Cathy Hayes)実験(1940s-50s): チンパンジー 家庭内でヒトの子供と同様に育て言語環境与える - 「パパ」「ママ」「カップ」等、4語程度の発音が可能と報告
発音不明瞭 - 意味の理解や文法的使用は確認されない
チンパンジー知能 = 高 ⇔ 音声言語習得には生理的限界 1971- LANA計画 (Rumbaugh et al): コンピューター制御語彙パネル
パネル上記号(語彙)を順番に選び命令文・質問文構成 → Ex. ゲザート地方のチンパンジー (松沢 2000) シロアリを穴から取るのに木を歯で噛んでスポンジ状のものを作る ≠ 道具を作るのは人間のみ 人間の1定義「人間は工具を使う動物である」 → 根拠: 他の動物は工具を作り、それで別なものを加工する技術を持たない[否定][証拠] 400万年前にアウストラロピテクス(脳容積600 cc)が打製石器を作る |
[ 霊長類化石 ]
|
人類: アフリカ起源の直立二足歩行をする霊長類の仲間
200万年より古い人類化石はアフリカからのみ 約600万年前にチンパンジーとの共通祖先から分かれ進化ケニアピテクス Kenyapithecus1500万年前 → ヒトやゴリラ等の共通祖先1997 日本ケニア合同隊: ケニアでほぼ全身骨格発見 → 大型類人猿
それまで、歯等部分的にしか見つかっていなかった ラマピテクス Ramapithecus: 1400-800万年前当初ヒトの祖先説 - 系統的に繋がらない(否定)形態 ≈ ヒト: 歯列弓アーチ型 切歯・犬歯小 臼歯歯冠低 1937 Lewis G Edward: New Deli北160 kmハリタリキングナールで発見ヒト科新属とし発表(当時アウストラロピテクスは認められていた) 1961 Leakey LSB: 2体目化石をKenya南西部フォートターナンで発見
上顎一部: 歯列アーチ型に近い ⇒ → ラマピテクス? (初期猿人は棚があるものがある) 1973 Turkeyのサンディールで下顎発見大戦中Ramapithecus?発見。Sivapithecusと報告 ⊂ ラマピテクス(後に) 1975 Hungary北東部ブタバニア山脈石炭層内で発見→ 当初ラマピテクスと分からない。現在も未断定 1977 2ダース程度のRamapithecus標本収集 ≈ 1500-1800万年前
ドミオピテクス: 森林生活(ゴリラの仲間) 300(500)万年前に猿人分化 → 第三紀にヒトと呼べる動物がいたはず 根拠 1. ルドルフ: 300万年前のヒト(?)顎 2. 上顎大臼歯: 磨耗 = アウストラロピテクス、形 = ラマピテクス → Africa人類起源説肯定されるか? 化石には化石となる条件が必要であり、そうとは言い切れない。化石の出来る場所・出来ない場所があり、Africaは化石形成条件の良い所で、化石の出来にくい場所でAfricaより早く人類出現があったかもしれない → 世界各地で調査発掘を行う必要 インド・パキスタン: 化石は全くでないが石器は出る1975 中国南西部 ラマピテクス発見(保存状態良)。アウストラロピテクス確認? ベトナム(戦争中): オラウータン発見 + ヒト歯(?)発見 イーストサイドストーリー East Sdie Storyグレート・リフト・バレー: プレートテクトニクスで1000万年前形成 – 西に山脈→ 東側: 気候大変動(降水量減少) → 森林から草原へ 初期のヒト科生物化石は、グレート・リフト・バレーの周辺または東側からしか発見されていない → 西側: 気候変動軽微: ゴリラ・チンパンジー・ボノボ(現在)二足歩行 H1 (旧). 草原化に伴い二足歩行化必要 H2 (新). 森林である程度の二足歩行能力獲得し草原に侵出 → ナックルウォーキング(チンパンジー) → ラミダス猿人 DNAによる系統分化推定ミトコンドリアDNA, mtDNA: 母性遺伝で組換考慮せず突然変異変化速度推定可能 – 時間的に一定割合で変異 構成が単純で分析しやすい ヒト-チンパンジーmtDNA組成差 ≈ 10% ⇒ 500万年前分化(ラミダス猿人?)猿人化石 Australopithecusaustralo = 南の, pithecus = 猿東アフリカ: エチオピア-モザンビーク(ビクトリア湖東側で多くの化石発見) 第三紀形成 - 代表的化石良く発見 (代表的土地 Oldvai Group)
東アフリカ発掘進むまで完全形発見されない 950万年前: サンブル・ホミノイド - ヒト(類人猿)祖先の可能性(未詳) 2000 ケニア・フランス合同チーム: ケニア北西部バリンゴで猿人骨発見
推定600万年前 =「千年紀の祖先millennium ancestor」と命名 |
2001 東アフリカ、エチオピア
520万-580万年前の猿人化石とみられる骨発見 サヘラントロプス Sahelanthropus2001 Beauvilain A, et al.: サヘラントロプス S. tchadensis
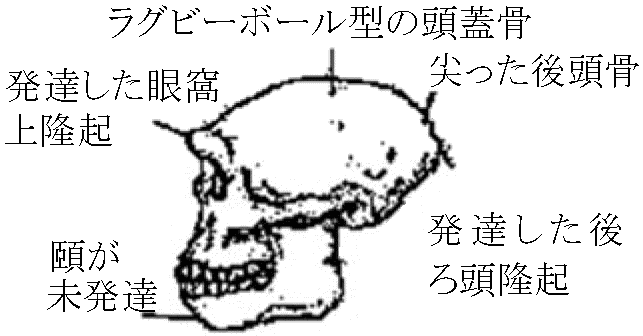
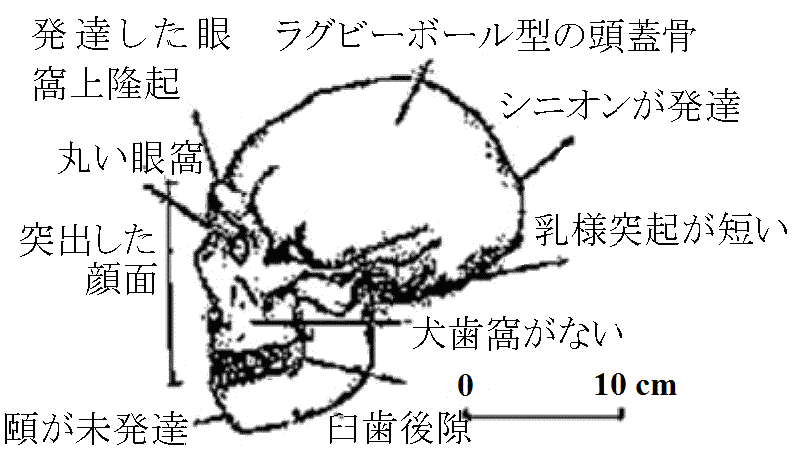
700-600万年前: 中央アフリカ(チャド)から頭骨 猿人の分化: 頑丈型と華奢型A. 頑丈型 (Paranthropus – Australopithecusから分ける)エチオピテクス猿人 (P. aethiopicus) A. robustus (syn. P. robustus): 南アフリカ 1947 Broom: 大人のAustralopithecus頭骨発見 → DartのAustralopithecusとは異なる → A. robustus 脳容積(cm³): 脳量がヒトとしての条件を決定するものではない
チンパンジー = 394 (<) オラウータン = 411 (<) 下顎・頬骨: 厚く張る 歯列: アーチ型 (ヒト = 丸い、サル = 角ばる、類人猿 = 中間的) 切歯: 犬歯小(顎水平移動可) Cp. アファール猿人: 大臼歯1.5倍、小臼歯2倍 歯磨耗度(すり減り方) (特に第1第2大臼歯): 大きく磨耗 – 動物食生活推定可 ⇒ 非常に固いもの摂食 – 果実、種子、茎、根等 + ×調理(×道具) 1959 Leakey: タンザニア、オルドバイ峡谷
Australopithecus boisei (P. boisei): 東アフリカ - 1.8-1 Mya 頑丈型絶滅 1995 アナム猿人 Australopithecus anamensis: 420-390年前 Ardipithecus (= 華奢型) 1922 Dart Raymond: Salmons, JosephineがDartを1922初期に訪問 1924 Dart R (1893-1988): スタークフォンテイン洞窟で頭骨化石 ⇒
タウングスで発見した古代ヒヒ(A. africanus)骨を調査 300-500万年前: A. afarensis (アファール猿人) ⊂ A. africanus 連続してハイデルベルグ人・シナントロプス等の人類進化証拠標本集まった事も発表理由 1936 Broom, Robert (1866-1951): Australopithecus transvaalensis脳 500 cc。歯列形状 ≈ ヒト = A. africanus 1947 Broom: 骨盤 pelvis 発見 - 直立二足歩行確実1972 Leakey & Leakey
260万年よりも下の層からヒト頭(脳量800 cm³)と石器発見
頭骨低部骨片: 大後頭孔は頭骨中央に位置 + 大腿骨形状 – 二足歩行
350万年前: 火山灰上に足跡化石発見
足跡: "土踏まず"があり、親指大きく発達 – 現代人と大差ない 3人: 足跡サイズ = 26, 21, 18 cm 26 cmと18 cmは寄り添うよう平行に並び21 cmは26 cmの上に重なる = 家族単位行動 → 二足歩行確実 1992 White & Suwa (諏訪 元)
ラミダス猿人 A. ramidus (syn. Australopithecus ramidus)
身長: ♂ 150 cm, ♀ 100 cm
→ チンパンジーと分かれて間もないヒトと考える
化石発掘層: 森林性種子動物化石多出 → サバンナ性動物化石皆無に近い - 森林生活? |
ホモ・ハビリス Homo habilis200-160万年前
脳容量500-600 cc + 顔面後退し小さくなる + 頬歯列やや小/前歯部大 1951 Leakey LSB et al.: 化石発見
Bet 2: 化石 + 石器 (Homo erectusのもの)
Bet 2: 礫石器発見 + 35 cm下から骨化石発見 = Homo habilisと命名
石器型を最低12種類に区別可能 → 各石器に異なる使用用途 石組住居確認。周辺に石器・動物骨・Australopithecus糞 - 食生活知る 1961 Leakey LSB: Zinjanthropus boiseiと命名した化石発見 + 石器
170-180万年前 石を打ちつけ割る → 石核石器(チョッパー): 手元に残る大型の石 + 剥片石器: 砕け散った細かな石
石核石器 – 斧・金槌として用いる → Africaには複数のAustralopithecusが260万年以前に存在 1967 Leakey Richard: Omo川流域調査
コービー・フォラで石器発見 (260万年 + α)
ケニア、トルカナ湖東岸で190万年前の頭蓋骨発見 1994 Carbonell Eudald et al.: グラン・ドリナ遺跡(スペイン)
80万年前 |
1994 Biddittu, Italo: ローマ南西59 kmで頭蓋骨発見
50-35万年前: H. erectusとH. heidelbergensisの中間的形質 →
時代(万年前)
┃ ラミダス猿人 400┃ ┃ 350┃ ┏ アファール猿人━━━━┓ 300┃ ┃ ┃ 250┃エチオピテクス猿人 ┓ アフリカヌス猿人 200┃ ┃ ┃ ┃ ┃ ┃ ┃ ホモ・ハビリス ┃ボイセイ猿人 ロブストス猿人 ┃ 150┃(絶滅) (絶滅) ┃ 図. 推定猿人 Australopithecus 系統 骨角歯文化(骨歯角文化) osteodontokeratic tool industry各所で確認 (≠ 石器文化, s.s.)資料統合 → Australopithecusが文化持ちヒトと断定(Dart) - 骨角歯文化
特徴: 工具を使っている 1925 Dart, et al. (+ 弟子達): マカパス洞窟で直立猿人相当の石器発見
別地にネアンデルタール人時代石器発見
Parapapio broomi (Parapapio 古ヒヒ, broomi 命名者)化石 = Australopithecus
脳容積750ccをサルとヒトとの境とする説が出たが根拠欠く
大量の骨はハイネナの仕業「Adam's ancestors」 ハイエナ説否定根拠: ≠ 洞窟に骨集積 ≠ 共食 (→ 洞窟にハイエナ骨) ≠ 骨食い残す 人骨 Australopithecus 発見 → 打撲跡よりAustralopithecus間に殺人行為(共食?)あったと考える |
|
1999 White and Suwa: 250万年前
A. garuhi: 近くから石器発見 → 初期原人の祖先? H. erectus, Syn. Pithecanthropus erectus (猿人と見なされていたため)A. Homo ergaster Groves et Mazak 1975: 180-140万年前, East Africa 初期原人 → 140万年前頃に後期原人のホモ・エレクトスへ進化 1980's 「トルカナ・ボーイ」: 160万年前の地層
アフリカ、ケニア、トルカナ湖畔
1999-2001 ジョージア、ドマニシで頭蓋骨・下顎骨発掘 原人(直立猿人): 身長175 cm – Australopithecusより現代人に近い 首から下は、ほぼ現代人と同じ 130-100万年前: Australopithecus africanus → 進化: Homo erectus
生存範囲広く、降雪地等でも生存確認 → 生存技術発達 アシュール型石器1.75 Mya: 出現 - 大型剥片を石器素材とするEx. ハンドアックス(石斧): 全体的に縦長で刃鋭い – 技術と熟練必要 アフリカ脱出Homo erectus (ジャワ原人)(Pithecanthropus erectus) 1891 デュボア(蘭): ヒトと類人猿の中間的なものを探す努力
トリニール川で歯化石一本発見。頭蓋骨の一部発見 = ヒトに似た一種の大型類人猿 (Sinanthropus pekinensis) 1923 北京周口店龍骨山(石灰岩)は名のとおり多量の動物化石出現 化石中に人骨混在判明 1927 Anderson: 周口店洞窟 本格調査開始: 哺乳類歯発見 = ヒト-サル中間発掘進め同様の歯発見 → S. pekinensisと命名 1929 下顎、更に頭蓋骨等発見 - ほぼ完全な頭骨出揃う1931 炉跡・石器発見 1937 発掘収量 – 計45体分の化石(太平洋戦争の混乱で紛失) ワイデンライヒ(米): 北京原人レプリカ保存。化石から報告書作成(解析進展)
→ 60-30万年前
洞窟: 大量の灰を発見 = 暖房(家) |
欧州 中国 北京原人 1907 独ハイデルベルグで発見 (60-20万年前)
形態的には現生人類と大差なし。脳容量 > 1200 cc
2003 Brown Peter et al.: インドネシア, フローレス島で発見 (10-5万年前) 身長 1 m程度, 脳容量 380 cc |
Homo sapiens neanderthalensis ネアンデルタール40-4万年前 (Syn. Homo neanderthalensis King 1864)1856 Fuhlrott JC: 独, セッツセルドルフ、Neander谷間の洞窟で発見
第四紀洪積世後期(20-5万年)
脳 :1300-1600 ml (最大1750 ml) 1911-1913 Boule, Marcellin (1861-1942, 仏, 解剖学): 半直立歩行 (否定)
「野蛮低脳。指さえ満足に動かせず、猫背で首を前に突き出し鈍重に歩く。現代欧州人とはかけ離れた種」 → 標本は重度関節炎患い猫背 ジブラルタル人: 既発掘ジブラルタル人頭骨再発見 ≡ ネアンデルタール人 ベルギーでの発見
1825 リエージュ近郊のアンジス洞窟で、子供の頭骨発掘
カットマーク(死体解体でできる傷跡)あるネアンデルタール人骨が20遺跡の砂地から散乱状態で発見 年齢40歳代: 右肩-腕萎縮し、右肘から先がない(生まれつき?)。右脚関節炎? 左目失明? 頭骨に治療痕 落盤事故で死亡? → 40歳まで存命したことは周囲の介護協力なしには考えにくい = 介護技術存在 1960 シャニダール洞窟: 6万年のネアンデルタール人化石発掘
化石周辺の土が軟らかく周囲に石並び遺体右向 = 埋葬 ↔ 反論: 「墓」という認識なかった (ガルゲット, 米 - ギャンブル支持, 英)
洞窟内で寝ている状態で自然死したものがそのまま残った 1984-88 イスラエル、カルメル山周辺洞窟遺跡群 ケバラ洞窟(5.5万年前) → ネアンデルタール人化石
1984: 舌骨(発音に使う筋肉がつく)完全形で発見 = 現代人と区別不可
(現在発見された中で)最古の埋葬 → ネアンデルタール人が新人(クロマニヨン人)に進化したとは考えにくい → 旧人(ネアンデルタール人)と新人(Homo sapiens)が共存していた |
1998: ネアンデル渓谷ネアンデルタール人上腕骨からmtDNA抽出 → 解析
ネアンデルタール人と現代人のDNA配列かなり相違 → 55-69万年必要 ムスティエ型石器最初フランス、ムスティエにて発見
60種類の型確認 = 石器製作技術進歩 + 主に剥片石器を用いたが使用目的により形の違う石器使用 狩猟技術に長じ、マンモス等の大きな獣をも狩る。火による調理 絶滅に関する仮説1. 無関係説 (エズラ・ズブロウ, 米, 先史学者)シュミレーション: 戦争がなくても、両集団の間の死亡率に2%の差があれば1000年程度で一方は絶滅 → ネアンデルタール人とホモ・サピエンスの両者間には「何もなかった」 2. 特殊化説: 分岐分類学a)原始形質: 原始形質祖先から(ほとんど)変化せず継承する形質
より古い人類であるホモ・エレクトゥスに見られる特徴がネアンデルタール人にも存続している形質 Ex. 脳容量の大きさが以前の人類よりも大きい c)固有派形質: ある集団にしか見られないその集団特有の形質Ex. ネアンデルタール人のイニオン上窩 ネアンデルタール人: 原始形質残しつつ固有派形質増加 → 保持型 → ある時期から環境適応困難
20-3万年前: ネアンデルタール人に大きな変化なし → 形質固定された 幼少時のくる病は死亡率高 = 化石にくる病推定させる骨格異常痕多 4. MVP説 3.6万年前: 人口がMVP下回った + 遺伝的多様性低下 + 絶滅雑種2010 Pääbo et al. 現代人DNAの1-4%はネアンデルタール人由来
ゲノム有: フランス人、中国人、パプアニューギニア人 |
イダルツ (ヘルト人) Homo sapiens idaltu White et al. 200316万年前(更新世) 東アフリカ1997 White, Suwa (諏訪), et al.: エチオピアのアファール低地で発見
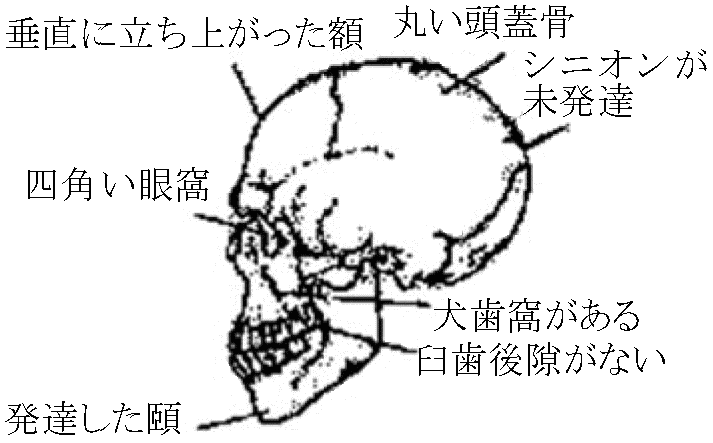
石器、死者への儀礼跡 クロマニヨン Homo sapiens sapiens洪積世後期(5-2万年) → 後期旧石器時代8-5万年前にアフリカ脱出 - 中東へ (DNA解析) 1868 南仏ドルドーニュ付近クロマニヨン洞窟 – ネアンデルタール発見直後
脳1300-1800 ml。下顎骨小さくなり頤がある。歯は現代人に似る。膝を伸ばした直立歩行 他所で記録された新人: ジャスラード人(南仏)、グリマルディ人(伊)、☛ 日本
中国: 柳江人、資陽人、周口人、上洞人(山頂洞人)
生活: 食物作る = 農業
2万年前壁画 - 馬・山羊・羊・野牛・鹿・カモシカ・人間・幾何学模様 シャテルペロン型石器 châtelperronian地域限定(スペイン北部-仏南部), 3.5-3.0万年前基本的作りはムスティエ型石器と同じ + 動物骨利用骨格器 + 高度装飾品(ビーズ、ペンダント) → 同じに出土 フランス、サン・セザール: 旧人。30000年前(ネアンデルタール人最新化石) シャテルペロン型石器が出土 → 旧人と新人との間に何らかの影響 (結論なく、今後の研究待たれる) 新人の起源a) 多地域進化説110万年前アフリカを脱出した原人起源 アジア、アフリカ、欧州各地で、原人が旧人に、旧人が新人に進化 (反論) ネアンデルタール人からクロマニヨン人に進化したとは考えにくい b) アフリカ起源説 ミトコンドリア・イブ仮説 (Willson A et al. 1987, 米, 分子遺伝学) アフリカ、アジア、欧州、ニューギニア、太平洋、各女性135人 mtDNA比較 |
仮定: 2-4%/100万年の変異確率。旧人-新人間遺伝的交流なし(= 調査された現代人遺伝子に旧人クラスのものが存在しない) → 20万年前アフリカ1女性mtDNA受継ぐ → 「アフリカ起源説」支持 → 新人: 20-10万年前アフリカで発生 → アフリカ脱出 → 個々の地域で新人 (反論) 20万年という年代推定には、最大100万年前までの誤差が起こりうる + 実際には、旧人と新人の間には交流があったと考えられる c) 東西二地域進化説 (馬場悠男)アジア: ジャワ原人 → 旧人・新人 (顔等の特徴が一貫して似通う) 欧州・アフリカ: クロマニヨン人がネアンデルタール人と交代 モンゴロイド起源は、原人か新人かは諸説がある(多地域進化説・アフリカ起源説)
┌────────────────┐
[北方モンゴロイド]←┤25000年前 12000年前?↓
東南アジア(中国) │ 北アメリカ
5000年前┌┴┐ │ 11000年前?↓
│ │ ├→3万年前 南アメリカ
↓ ↓ │ 1500年前┌→ハワイ
[南方モンゴロイド]─────┼→イースター島
スンダランド┐ 4000年前 │ 1500年前
6-5万年前↓ 1000年前└→ニュージーランド
オーストラリア
スンダランド年代分け各地へ移動6-5万年前: サルフランド(当時オーストラリア、ニューギニア、タスマニアが陸続きの大陸)に移動
数万年間移動は断続的に起こった 25000年前: 北緯60°を越えシベリアに至る 技術革新 = 火・衣服・細石刃(カミソリ状の石器) 12000年前: 北極海に達する → ベーリング海渡りアメリカ大陸へ = アメリカ先住民 12000年以上古い遺跡がアメリカ大陸で発見 → 民族等は不明
縄文人はこの時期に移入した人々と考えられる 2300年前: 渡来系弥生人はこの時期に移入した人々と考えられる |
日本: (弱)酸性土壌多 - 骨溶かしやすく化石残りにくい
原人 (archanthropine, early man)60万年前から日本にヒトと呼べる動物がいた1905 マンロー(英): 神奈川県酒匂川流域で礫器 1949 群馬県岩宿遺跡 相沢忠洋(行商人、考古学マニア)発見
10万年前の関東ローム層から石器発見 石器 – 中国周口店(北京原人)石器と似る 1984 宮城県古川市馬場壇A遺跡: 15万年前 → 石器出土1993 宮城県筑館町高森遺跡: 50万年前 → 石器出土 1994 宮城県筑館町上高森遺跡。60万年前 石器出土: 石器発見/人骨出ない → 日本土壌の特性 1968-74 沖縄県本島具志頭村港川採石場石灰岩割目
港川人 (18000-16000年前) – 完全に近い人骨4体
男子153 cm、女子144 cm – 縄文人身長に近い 人骨出土1959-61: 静岡県三ヶ日町(現浜松市北区)石灰岩採石場三ヶ日人 (9000年前 - 2001年14C測定) 1962 沖縄県山下町第一洞穴遺跡: 前旧石器・焼土(炉跡)・焼礫
4.0-3.6万年前 = 旧石器時代
14000年-10800年前(旧石器時代) - 14C 聖岳洞穴人(聖岳人)・黒曜石製旧石器 – 新人(後否定) 1975 愛知県豊橋市牛川町石灰岩採石場牛川人 – 人骨(旧人)ではない? 1950 栃木県葛生町(現佐野市)石灰岩地帯
葛生人: 上腕骨,大腿骨,下顎破片 1932 直良信夫 兵庫県明石市海岸にて原人人骨?拾う
1945 空襲で化石消失: 松村瞭助(東大) 直良人骨写真と石膏模型保存
ジャワ原人と同段階 → Nipponanthropus akashiensisと命名 石膏模型コンピューター人骨比較解析 → 縄文時代以降の新人 縄文人 (Jomon man)1952 関東ローム層集団研究 - 動植物遺体(化石)が石器とともに出土しない25000年前 立川ローム層 (17000: 第一黒色帯, 25000: 第二黒色帯) 広く同一年代の地層 - 後期旧石器 ⇒ 日本全土に人類 1962 野尻湖: (長野): 5万-3万3000年前の動物化石群
ナウマンゾウ臼歯 (後に全身、オオツノシカ化石)
2万年前(氷河期最盛期): 日本の大部分森林 - 北海道: ツンドラ 気候: 沖積層世(沖積世) → 温暖期: 森林(気候帯)北上 5000年前: 気温↓ 3500年前: 気温↓↓↓(Santorini火山大噴火) 遺跡上野原遺跡(鹿児島): 早期 → 最古級集落跡加曾利貝塚(千葉): 中期-後期 → 最大級貝塚 大森貝塚(東京): 晩期 菜畑遺跡(佐賀)・板付遺跡(福岡): 晩期 → 水田・水路跡 人口: 最盛期 = 30万人 (奈良時代 500万、室町時代1200万)
東北地方 46000 vs 近畿中国地方 4000 生活: 竪穴(式)住居 (pit dwelling) 家 = 4-6本の柱(50-70 cmの穴) + 茅葺屋根 = (半)定住生活 (大船遺跡) 主食料: ドングリ類、獣、魚介類
縄文クッキー: アク抜した堅果(ドングリ等)の粉を焼き作る – 貯蔵法発達 土葬: 大人と子供埋葬地違う(大人: 住居離れる - 子供: 入口, 七五三起源?) 縄文土器 Jomon earthenware vessel: 13000年前 – 世界的に最古
縄目模様独特 – 呪術的意味?
1975-1985 発掘作業 「縄文のタイムカプセル」 ヒョウタン果皮 - 人為的加工跡 照葉樹林だがクルミ・クリ多量出土 = 定住生活開始? → 人里植物出現 1980-1985 発掘調査: 5.5 Kya 縄文前期 = 漆櫛、漆鉢、漆盆
2011: 12.6 Kya ウルシ枝発見 - 世界最古の漆使用 最大500人規模集落 → 1500年続いた巨大集落 長さ30 m程の長屋状住居跡。直径90 cmのクリ材柱(栽培可能性) 北海道産黒曜石出土 [縄文人 → アイヌ民族] |
ヒスイ(翡翠)出土: 新潟県糸魚川が日本唯一の産地 丸木舟 = 「海の道」交易 朝鮮半島の縄文時代同時代遺跡から日本産黒曜石出土 千葉大坪貝塚: 縄文時代中期 - ヒョウタン岐阜ツルネ遺跡: 縄文時代中期 - 炭化したマメ科(エンドウかダイズ)種子 埼玉上野遺跡: 縄文時代中期 - オオムギ?炭化物 長野伴野原遺跡: 縄文時代中期後半 - パン状炭化物と小炭化物(マメ子葉?) 長野荒神山遺跡・大石遺跡: 縄文時代中期 - エゴマ(作物) ヒト
[採集] ━━━━━━━━━━━━━┳━━>
[クリ-クルミ林発生] 森 稲渡来 ━━━━━━┻━━━━━━> 稲作文化発達 漆器、精巧土器(遮光器土偶出土)、ヒスイ製玉 是川遺跡(青森) 縄文時代晩期中心
低湿地: 漆製品(漆塗弓・櫛、腕輪、容器等) 弥生人 (Yayoi man)弥生時代 = 弥生式土器使用時代(BC5-4c–AD3c): 特徴 = 稲作、金属器言語: 基礎言語 = 南方系言語 &rarr: アルタイ系言語の影響加わる 1957 大野: 縄文時代 - 南方系言語(ポリネシア語族様音韻組織)移入
弥生文化伝来 = 朝鮮南部言語(アルタイ語的文法体系・母音調和)流入 + 大陸系言語(アルタイ系言語)加わる = 日本語 1976 西田: 上代日本語とチベット・ビルマ語比較日本語基盤 = チベット・ビルマ語系言語 1980 大野: タミル語が照葉樹林文化と共に伝播 - 日本語形成に関係(対立仮説ではないと思う) 日本語起源は縄文時代まで遡る必要 遺跡菜畑遺跡(佐賀)・板付遺跡(福岡): 早期, 水田・水路跡(縄文晩期より続く)唐古・鍵遺跡(奈良): 中期, 奈良盆地最大の中期農耕集落 吉野ヶ里遺跡(佐賀): BC100-AD0, 最大の環濠集落(高床式倉庫・物見櫓)
= 邪馬台国?(卑弥呼の時代と合わない) 砂沢遺跡・垂柳遺跡(青森): BC-AD, 田跡 原の辻遺跡(長崎): AD1c, 3重環濠、最古の船着場、中国貨幣(1c鋳造) 4000年前: 寒冷化 = 現在の気候と概ね同じ 縄文人人口最盛期30万人が75千人に減る 化石産出: 九州中心西北九州型: 長崎、佐賀南部 = 縄文人人骨に近い → 縄文人起源 北部九州型: 山口、福岡、佐賀北部 = 身長163 cm、顔立面長 – 形態的に縄文人と大きく異なる
→ 断続的変化で縄文人が北部九州型弥生人となったとは考えにくい
混血は起こる → 現代へ BC 2 c-AD 1c: 出雲銅剣文化
1984 島根荒神谷遺跡 - 銅剣358本 (出雲式銅剣: 長さ52 cmと長大)
1985 銅鐸6個、銅矛16本 - 初(銅剣 + 銅鐸 + 銅矛 組合わせ)
1996 島根加茂岩倉遺跡: 銅鐸39個 → 出雲が青銅器祭祀一大中心地
→ 稲作伝播速度は極めて早い ☛ 稲作伝播
AD57: 倭 奴国王が後漢朝貢 - 光武帝から金印(印綬) - 志賀島で発見 環濠集落: 集落を濠で防備 Ex. 吉野ヶ里遺跡(佐賀)・唐子-鍵遺跡(奈良)・原の辻遺跡(壱岐)・池上曽根遺跡()・板付遺跡(大阪) 高地性集落: 小高い丘の上に作られた集落 → 低地性集落(従来): 平野部に作られた稲作目的の村
物見が目的 → 瀬戸内海沿岸、近畿地方に多 → 大和大乱説: 『魏志倭人伝』「倭国乱れ、相攻伐すること歴年」 → 集落防御目的 AD 4c: 騎馬民族日本征服説(江上 1967) → 反論も多い古墳巨大化 + 急激に戦闘的になる – 騎馬民族(Ex. 匈奴)侵入? → 北東アジア蹂躙した騎馬民族が日本に入り社会中枢を占めた 渡来人の存在は疑いない Ex. 墓制の多様化 → 朝鮮半島の影響と支配層(豪族)の搭乗
弥生式土器・銅鐸 |
☛ 本州年代対応 北海道 |
|
|---|---|
|
石鏃: 鋭い先頭部と形態様々な基部からなる扁平で小型の石器 - 主に矢尻
1.5万年前: 出現 - 弥生文化では武器としても使用 基部形態は時代と共に変化 - 北大構内でも発掘 旧石器(先土器)時代50万年前: 上川郡清水町美蔓西遺跡(否定 ∵ 旧石器捏造判明)20万年前: 新十津川町総進不動坂遺跡(否定 ∵ 旧石器捏造判明) 12万年前以前: 前期旧石器時代 1999 長崎: 両面加工尖頭器、採土スクレイパー - 洞爺火山灰層(10万年前)下位 5万年前: 石器(斜軸尖頭器)1998 長崎: 石狩川右岸ローム層露頭(樺戸町新十津川総進) 2万年前: 北海道は海水面下降し大陸と陸続き → マンモスや人類渡来
マンモス化石: 襟裳岬、夕張市、空知管内由仁町 打製石器は石刃、細石刃、尖頭器、石錐等 → 魚介類、オオツノジカ(更新世北方系動物)、野牛、トナカイ等を獲っていた
網走管内白滝村から当時(2万数千年前-縄文時代)の遺物が豊富に出土 縄文時代(文化), 15-2.4 kya打製石器 → 磨製石器初期: 海水面現在より3-5 m↑ (縄文海進) = 海: 釧路・網走・石狩・苫小牧近辺
道南: 椴法華式に代表される尖底土器多く出土(本州東北地方と共通) 石刃鏃 flint arrowhead: 石を打ち欠いた石刃で作った鏃 石刃鏃文化縄文早期中-後葉期: 石狩より北部で出土日本: 北海道だけで出土 (十勝・釧路・網走方面に多)
大規模遺跡: 道東北部沿岸部に集中 - 漁労重要 遺跡垣ノ島遺跡 Kakinosihma Site (函館市臼尻町)縄文人: 縄文時代早期前半-後期後半(約9-3千年前) - 地域の中心的ムラ 集落遺跡: 太平洋に面す高台 = 渡島半島東岸、垣ノ島川左岸の丘陵地 5000BC 居住域-墓域分離 = 日常生活だけでなく精神世界示す遺構・遺物
墓副葬品 = 儀礼道具 Ex. (幼児)足形付土版、青龍刀形石器や石棒、漆塗り注口土器やヒスイ製装飾品等 (2011年2月7日 国指定史跡 指定面積 92749 m2) 縄文時代前期大規模貝塚を中心とする集落遺跡 ☛ 縄文の森
噴火湾に面した舌状丘陵上(保存状態良好) 縄文前期-晩期自然環境、生活生業、人類学的形質や葬法等知る上で重要 |
入江貝塚: 縄文前-後期の大規模貝塚伴う集落遺跡
厚さ3 mに及ぶ貝層から多数の人骨・遺物出土 高砂貝塚: 縄文後期 + 縄文晩期土坑墓群も確認。多数の人骨と副葬品出土鷲ノ木遺跡(森町): 現海岸線から1 km内陸の舌状丘陵上 + 保存状態良好 縄文時代後期前半の環状列石と竪穴墓域 環状列石: 中央4 × 2.5 m配石と二重列石で構成、外周37 × 34 m 円形 道内最大規模 竪穴墓域: 環状列石に隣接 11.5 × 9 m ほぼ円形 大小11基の土坑墓 北日本環状列石・墓制・祭祀等の精神世界 + 北海道-北東北交流
100棟以上の竪穴住居跡 (> 深さ2 m 大型竪穴特徴) + 盛土遺構、土坑群等 大量土器・石器の他、クジラ、オットセイ (+ 銛 harpoon head)、クリ、ヒエ等動植物遺体出土 → 生活・生業(漁業 + 農業)知る上で重要 竪穴住居 (pit dwellings): 縄文時代住居形態。地面を円形・方形に掘り床とし、柱を立て骨組作り、その上に土やヨシ等で屋根をふいた建物で、床には炉が設けられる。大船遺跡の住居は大規模で、長さ8-11 m、深さ2 m以上の住居がある。炉は、床そのものを火床としたものから土器を埋めたもの、石を囲んだ炉へと変化が見られる。本復元住居は、発掘調査の成果を元に、クリの木を使い竪穴住居の骨組を再現した。(2018/7/11) 後期・晩期道東では遺跡は、殆ど見られなくなるアスファルトの道
磨光B遺跡(南茅部町): アスファルト塊 = 矢尻・矢柄接着に使用(加工技術必要) 職人集団存在の可能性 + 貿易拠点の可能性 環状列石 (☛ 忍路環状列跡)続縄文文化本州弥生時代に、北海道に鉄器伝わるが稲作農耕行われず、権力者も出現せず、縄文文化の伝統を残した文化続く前期 = 本州弥生時代
後北式文化: 道東、道北、道央 - 後期には東北地方まで伝播
後北式土器が全道に広がる Ex. 余市町フゴッペ洞窟
秋田県能代市寒川II遺跡 = 後北式文化 → 青森県下北半島-八戸、上北地方、宮城県、秋田県 「港大照寺遺跡」跡地埋蔵文化財埋蔵地 - 昭和44, 45, 47年に発掘調査実施縄文晩期の日本海沿岸で初と言われる積石墳墓群や石器・土器片多数発見 この時期の太平洋沿岸以外の墳墓等となる港大照寺遺跡の発掘は、北海道の墳墓の構造や葬法を知る上で貴重 (平成30年10月 蘭越町 蘭越町教育委員会) |
|
個人(心理学) ⇔ 社会(社会学) ⇔ 文化(人類学): 相互作用 = 文化人類学 文化の過程 culture process 文化の統合 culture integration
研究上の障壁(全ての統合科学に言える): 「人よ、汝自身を知れ」 ある特定社会の生活様式を1つの全体として研究し明らかにできる問題がある 文化型: 行動の標準 → 文化: 個々の社会が持つ生活様式。「習得された行動と行動の総合体であり、その構成要素がある1つの社会のメンバーによって分有され伝達されるもの (Linton 1945)」 社会により生活様式が異なる = 民俗学 民族生態: 人類生態系を民族集団単位で比較し、民族間相似性・相違性を知り人類の生態的特徴を知る多方面からの調査必要。至極一般的な(ありふれた)ものを追求する必要 [他国文化理解 ≠ 未発展文化を強制的に変革させ先進的にする] 未開社会: 文字のない社会 (軽蔑的な意味ではない) 世界中の文盲率は40%(35%) → 文盲 ≠ 野蛮 primitive → 厳しい生活に対する adaptation ともいえる Ex. エスキモー・ラップ人・ブッシュマン・マサイ・アボリジーン (Ref. ウォンランド「古代社会」) 人種 race (ethic group)人間の遺伝的特徴によりなされる分類アジア系(黄色人種): 皮膚黄-銅色、毛髪黒色直毛 Ex. アジア諸地域、アメリカ先住民 欧州系(白色人種): 皮膚白色、毛髪金髪-黒色波打つ、伸長高く高い鼻Ex. 欧州、アラブ、インド アフリカ系(黒色人種): 皮膚黒色、毛髪黒色巻毛状Ex. 中央-南アフリカ諸地域 その他(アジア系としてもよい) Ex. アボリジニー、ポリネシア民族 ethic group (race)文化的(言語・宗教・習慣・社会構造等)な分類 → 人種・民族差別(主要)言語 language
中国語 Chinese: 使用人口最多 宗教 religion6000年前: 高温期 = 農耕発展 - 多神教発生5000年前: 気温低下 + 乾燥 → アジア・アフリカに乾燥地帯発達
砂漠化による神々の脱落 → 一神教発生
3200年前: カナン定住 - ラクダ家畜化 受肉 inculturation: 神が人の形をとり現れること Ex. イエス、ブッダ • キリスト教 Christianity: カトリック(旧教): バチカン中心。ラテン、中・南アメリカ プロテスタント(新教): ゲルマン民族、北アメリカ ギリシア正教: ギリシア、ロシア、東欧 原罪 (original sin, peccatum originale L) アダムとイヴから受け継がれた罪 (キリスト教西方教会) • 仏教 Buddhism:上座部仏教(南伝仏教): インドシナ半島、スリランカ 大乗仏教(北伝仏教): 日本、朝鮮、中国、ベトナム |
♠ キリスト教 - 仏教
西と東
直線的世界観(有限) - 円環的世界観(無限) 天地創造-終末 ×受肉 スンニ派(正統派): イスラム教徒の90%シーア派: イラン中心 • ヒンズー教(インド) バラモン教 文化 culture
↓ 個別的文化 社会構造人間生態系 human ecosystem
人類は自然界で生態系の内容を人為的に変え、その変化は農耕・牧畜により促進され、更に近代産業革命により爆発的な変化をしている 環境: 自然と関わる方法の他生物との違い
人為環境(二次的環境) man-made (secondary) environmentを介在させ地球上の殆ど全環境に適応 I 乾燥地帯と砂漠 採集: ブッシュマン オーストラリア原住民
遊牧: ベドイン トウアレグ 単純農耕: アマゾン住人 複雑農耕: ニューギニア住人 III 熱帯灌木 採集: グランチャコ・インディアン 狩猟: バンドー
牧畜: ベンバ
狩猟: ハッザ
狩猟: タスマニア島人 プシエドモス人 (東欧のマンモスハンター)
牧畜: バルカン人
遊牧: ベルベル
牧畜: モンゴル人
遊牧: ブリアート モンゴル人 IX ツンドラ 狩猟: エスキモー 遊牧: ラップ 環境区分と生活経済とはあまり相関性がない。しかし、人間生活にとって環境が重要な要因であることに変わりはない。例えば、土に合わせた作物の選択、水供給を考慮した住居の決定等である |
|
諸民族の文化の特質を歴史的、あるいは他文化と比較し研究 ≈ 文化人類学 特質: 生活様態・文化 Ex. 言語、宗教、社会制度、法制、芸術、技術 1958 岡 「日本民族の起源」: 種族文化複合 = 移入文化は複数 → 重層融合
(1) 母系的・秘密結社的・芋栽培 = 狩猟民文化 (証拠不足) 中国江南地方から4-5世紀に渡来
(5) 父権的・「ウジ」氏族的 = 支配者文化 (アルタイ系言語種族文化) 照葉樹林文化 evergreen broad-leaved forest culture1966 中尾: 照葉樹林帯に共通文化 (☛ 照葉樹林)
Ex. 水晒(灰汁抜)、茶葉加工飲用、漆器製造、製糸(絹)、柑橘類・シソ栽培利用、酒(糀使用) + 稲
→ 照葉樹林分布域に伝播 → 照葉樹林農耕文化(複合文化)
重要な文化共通点: 雑穀類栽培焼畑農業
イモ類(サトイモ、ナガイモ)、アワ、ヒエ、シコクビエ、モロコシ、オカボ
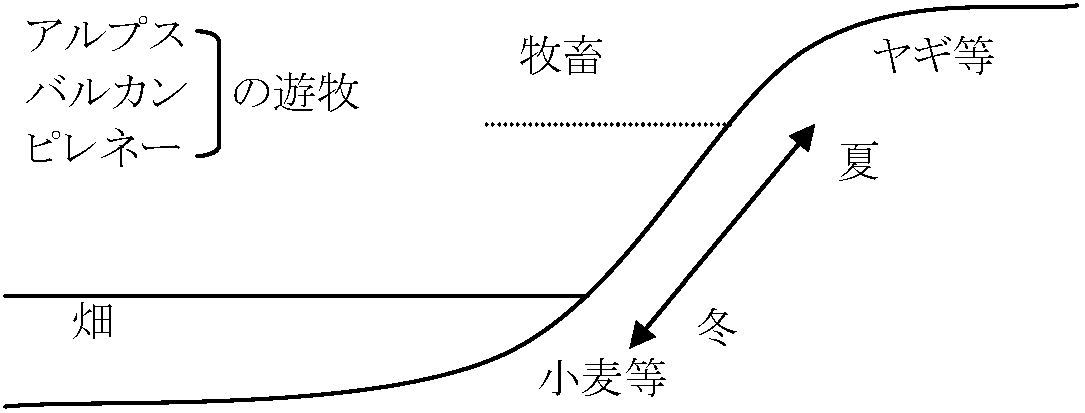
稲作文化に先行する文化 = 日本稲作文化の母体 儀礼食: イモ → イモ・餅 → 餅 (変化: 畑作(焼畑) → 稲作) 東亜半月弧 East Asia Crescent: 雲南省かその周辺が中心 → 文化の伝播イネ・ダイズ・アズキ・ヒエ・ソバの起源地
+ 茶・絹・納豆・蒟蒻・漆・紫蘇・味噌・麹・鵜飼等々 共通文化要素多 = 文化中心 (仮説) + 多民族地域 7-6 Kya: 農耕成立 1764 辰州府誌: ミャオ族の記録は鳳凰庁志と同ヤオ族: 焼畑(雑穀栽培) + 茶・漆植栽 1824 鳳凰庁志: 湖南省西部山地ミャオ族記載 - 焼畑記録山傾斜地で焼畑農耕(= 刀耕火種): アワ、シコクビエ、モロコシ、ハトムギ ⇒ ミャオ族・ヤオ族: 陸稲栽培記録なし(辰州府誌・鳳凰庁志)1921 Hutton: アッサム地域北東部(東亜半月弧東部) ナガ族 - ナガランド
セマ・ナガ族: 焼畑2年栽培10-12年休閑 陸稲依存度は地域差 (∵ 気候) Ex. 高地はハトムギ主体 2年目耕地: 雑穀、豆類 (∴ 雑穀栽培型焼畑) 1937 Mills: レングマ・ナガ族 → サトイモ主 - 死者への供物1938 Stübel: ヤオ族焼畑の記録 - 瑤山: 焼畑 = 3年栽培10年休閑 (主)オカボ、サツマイモ、トウモロコシ + 雑穀類 植栽: 油茶樹 + シュロ(繊維) + 周囲: 野菜類、豆類、サトイモ
計竹杭村: 加えてハトムギ栽培の記録 = 照葉樹林における伝統的な生活 ≈ 雑穀-根菜型 1939 Fürer-Haimendorf: ナガランド東北部コニャク・ナガ族焼畑(調査1938)2年栽培10-40年休閑 - 陸稲、雑穀、サトイモ ↔ 水稲はない ⇒ サトイモが古くからの主食 + イネは新しい(説)1981 武村「ヤオ族の歴史と文化」
湖南省照葉樹林帯起源 → 広東省北部 → 福建・浙江 / メコン地域 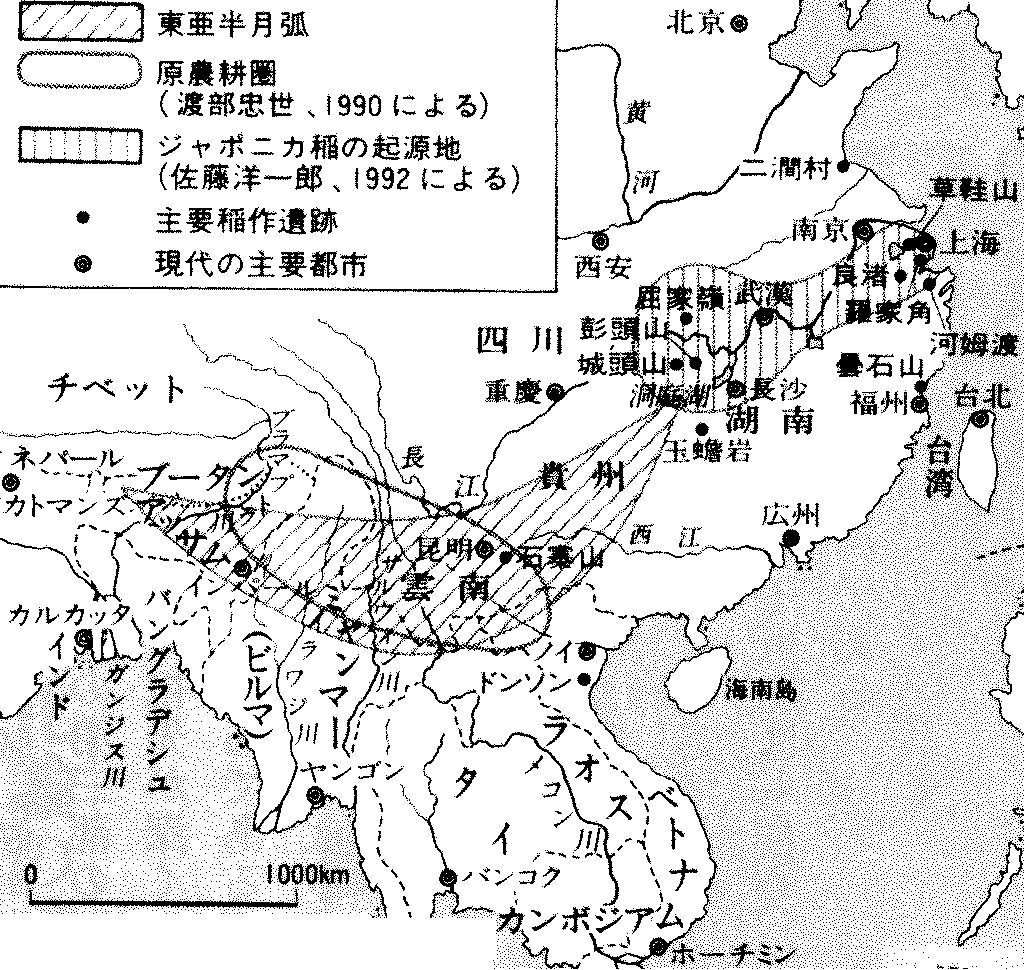 図. 東亜半月弧と原農耕圏とジャポニカ稲の起源地 ブータン東部・アッサム中東部-北ビルマ-雲南-貴州省中南部-湖南省西南部 本来照葉樹林(現存はブータン・アッサム) 表. 照葉樹林文化の農耕方式の発展段階 (中尾 1967)
1. 照葉樹林採集・半栽培文化(プレ農耕段階) - 灰汁・毒抜き重要 縄文時代前・中期(BC4000-BC2000) (a) 叩き晒し法 water bleacing (毒抜き detoxification - protocol)
日本: ヒガンバナ = 毒抜きし食用 Ex. ワラビ粉、野生イモ、クズ 水晒しの方が加熱よりも大量処理が可能: 様々植物の灰汁毒抜きに応用Ex. 落葉広葉樹林: トチノキ、ナラ類、シイ類 (b) 加熱処理 boiled bleaching: 本処理による毒抜きは照葉樹林帯では少1796 「巡島日記」: 八丈島 - Arisaema spp.を加熱毒抜き後に食べる |
伊豆諸島利島では叩き晒し法で毒抜き
ヒマラヤ中腹でもArisaema何種か加熱による毒抜き後に食用 水晒し汁抜きは熱帯地域では少 ⇔ 照葉樹林 半栽培植物 - 保護・管理・育成(≠ 栽培)Ex. ヒガンバナ、ワラビ、マムシグサ、ウバユリ、クズ、カジノキ、ウルシ 人里型 ruderal: タイの野生(推定)サトイモ(2倍体) = 実は選別受ける ∵ 野生型と遺伝子異なる 日本: コウボウイモ(3倍体) = 野生型サトイモ(種子繁殖不可) 縄文時代早期末-中期早に渡来? → 半栽培 Ex. 鳥浜貝塚
食料安定供給: 堅果類・球根類?・貝類・淡水魚・狩猟(狩猟)
= 原初的農耕 incipient agriculture 縄文時代後期-晩期: 徐々に農耕のウェイト高 Ex. 西日本照葉樹林帯 東南アジア熱帯モンスーン林地帯(照葉樹林帯南側) タロイモ、ヤムイモ - 栽培化 (更に南: バナナ - 栽培化)
↔ 東日本ナラ林帯 - 採集社会継続
照葉樹林文化に影響 = 雑穀-根菜型作物複合体形成 縄文時代末(弥生時代初頭)頃
1966 可児「鵜飼」: 鵜飼は稲作と共に伝播 ⇒ インドアッサム-ブータン: (家蚕蛾科 vs 天蚕蛾科) × (飼育種 vs 野生種) Bombyx mori = 家蚕蛾科完全飼育種、変種有 絹生産の起源は東亜半月弧? 漆器 ☛ 漆紙文書
1980-85 調査: 縄文前期(5.5 Kya) 鳥浜貝塚 ☛ 漆器
酒: 発酵酒 = 照葉樹林文化圏特有
サトイモ、ヤマノイモから繋がる粘性高い食物への嗜好性 → 6C中 宗懍(梁)「荊楚歳時記」: 儀礼食としてチマキ記載 神話・儀礼・習俗1973 大林 「稲作の神話」 作物起源神話: 死体各部分から作物(死体化成神話)
日本: 五穀 = 稲 + 粟 + 小豆 + 麦 + 大豆 = 穀物 記紀の五穀 = 本来は穀物ではない? → 雑穀栽培型農耕文化 山海経: 戦国-秦朝・漢代(BC4C-AD3C)に付加執筆され成立 - 最古地誌 后稷(農耕の女神)が葬られた所に雑穀生じる 神婚神話・国産神話・洪水神話
イザナギ・イザナミ神話に類似した話は東南アジアに広がる ⇒ 昔話(Ex. 竹取物語、羽衣伝説)にも照葉樹林帯で共通点 焼畑文化の影響も見える 歌垣: 共通点 = 風土記内容 + 口上 (+ 妻問)お月見(十五夜)を祝う風習 儀礼: 日本を含む東南アジアに共通点多
表. 照葉樹林帯8種族(コニャク・ナガ、セマ・ナガ、カテン、ヤオ、ミャオ、ブヌン、ルカイ)の焼畑栽培作物。*新大陸起源. 主(要)、副(次) (佐々木 1982)
日本の焼畑: 伝統的焼畑輪作72村(作付率, % = 作付村/全村)
主 副
作物 アワ中心に多様な穀類 + タロイモ中心としたイモ類
雑穀-根菜型が広く分布 ソバ、ヒエ: 雲南山地以東 ハトムギ: 照葉樹林帯辺縁部に集中
アワ(83)、ソバ(81)、ダイズ(76)、アズキ(76)、ヒエ(57)、ムギ(21)、サトイモ(19)、サツマ(15) + 北方ナラ帯文化 Ex. 九州山地部: ムギ、(エンバク、カブ、ダイコン) ⇒ 日本文化 ≠ 稲作文化 = 非稲作文化 (焼畑文化の名残多) + 地域差
イネは雑穀の一種 ⇔ 稲作: 単作(随伴種がない) ≠ 作物複合体 雑穀の中から単作作物として選ばれた? 耕地条件[焼畑] ━ (テラス化・集約化) ━┳━ [水陸両用の天水田] 常畑 ━━━┫ ┃ 畦畔構築・耕地水平化・灌漑施設築造 ┃(水理条件改善) ┏ 栽培条件改善 ⇒ [畦畔持つ水田] ┃ ⇓ 水稲単作 ┣ [収穫安定] ⇐ 水稲品種群形成 ┗━━━━━━━━━┛ 栽培条件
雑穀混播・散播 収穫不安定 → 雑穀混作・条播 収穫やや安定
天水田(佐々木造語): 水利を天水(雨水・雪解け水等)に依存した水田
インドビハール州、中部ネパール等 - 原初的天水田 → 収穫・脱穀・調整技術未発達 6-5 Kya 青蓮崗文化(淮河、長江): 上海付近崧沢、江蘇省草鞋山 - イネ出土5 Kya 屈家嶺文化(長江中流)、良渚文化(下流) - 多量のイネ
屈家嶺文化: 紅焼土(土壁築造用の焼けた土塊)から大量の稲籾
磨製石器、尖底土器 = 農機具、調理器具 ⇒ 水田稲作農耕といえる段階 |
☛ 日本史
|
宇宙考古学 space archaeology: 衛星から地表の見えない遺跡を探索
古代文明発見 Ex. エジプト・中国古代都市遺跡を衛星データから特定 概説 (中国本土との関係)匈奴: モンゴル系(モンゴロイド)説有力、トルコ民族説、イラン民族説 BC4c終 発現(モンゴル中央部)
BC 221-BC 206 泰: 始皇帝 → 万里の長城・蒙恬派遣 武帝: 西方(月氏)へ張寫派遣するが失敗 BC209 冒頓単干(単干: 王称号)冒頓が部族(モンゴリア)統一 → 月氏を西方へ追放 AD1c 勢力衰退48 南匈奴 漢吸収 91 北匈奴 鮮卑のため中央アジアへ移動 鮮卑: モンゴル系説(有力) vs トルコ系説 1c 進出 - 壇石愧, モンゴル高原から中国進出
304-439 五胡十六時代
9c 進出_____BC 2c: 匈奴に支配される 5C 社崙 丘豆伐可汗(可汗 = 王位: 小可汗、葉護 = 諸侯) 突厥 (Türk): 匈奴の末裔。原トルコ的特徴有 トルコ系(アルタイ地方原住)
西丁零起源 552 柔然から独立: モンゴル高原北西部に部族連合である突厥可汗国建国
長男 土門(=伊利可汗/土門可汗 552-552) 次男 室點蜜
→ 部族反乱等で勢力衰退 657 西突厥: 西可汗阿史那賀魯が唐の捕虜 - 唐に服従 ≈741 西突厥滅亡 回鶻: 現ソ連、ウイグル自治区の民族祖先。トルコ系 745(or 744) 突厥滅ぼす
618-907 唐 当時遊牧国家も乱れ、中国本土への侵入少 |
クトゥルクビルガ(懐仁)可汗がウイグル統一 キルギス: エニセイ・オビ川上流840 ウイグル滅ぼす 契丹: 内蒙古東部地域 915 民族統一: 国名 遼、邪律阿保機(太祖) 960-1279 宋: 趙匡胤 1125 金に滅ぼされる↓ 西方へ逃亡 1132 西遼(カラキタイ) (-1211 チンギスカンに占領される)女真(直): 半農半牧。ツングース系 1115 阿骨打(太祖)。国名 金 モンゴル (1279-1368): モンゴル系 成吉思汗(チンギス=ハン) Chinggis-Khan (1162?-1227) 1206 モンゴル帝国 = 民族統一 (商人厚遇) 1235 カラコルム Karakorum - 首都 忽必烈(フビライ) Kubla Khan (1215-1294): 第5代皇帝、成吉思汗孫1264 大都(現北京)遷都 1271 国号改訂 大元 yuan = モンゴル帝国後裔 (東アジア-北アジア)
北元: 明に追われ北に逃亡(モンゴル高原)
1243-1502 キプチャク (金帳, Kipchak) 亀石 turtle rock - 碑文の台座 1368 明に滅ぼされるティムール帝国 (Timurid Empire) 1370-1507: トルコ化モンゴル系 ティムール(Taimur, 1336-1405)建国: 中央アジア 明 (Ming) 1368-1644: モンゴル系1368 元滅ぼす 清 (Qing) 1636-1912: 女真の末裔1616 ヌルハチ(太祖) 後金建国/ツングースとモンゴルの統一 1636 清建国ジュンガル王国: モンゴル系 1634-1755: オイラート族の末裔 / 新彊 1911 外蒙古、モンゴル独立宣言 1912 中華民国成立 1922 モンゴル人民共和国(反清運動) 1949 中華人民共和国成立 |
| 遊牧論 | |
|---|---|
|
Def. 遊牧: 動物が一定ルートをとらずに広い範囲を日数をかけ移動すること Def. 遊牧民: 家族全員が家畜と一所に移動 Ex. 蒙古: 甘粛省・青海省・チベット北部等。ベトウィン、ベルベル族、モンゴリア人: 牧畜 = 経済生活全てあるいは大部分に牧畜産物利用 家畜・食料・衣服・住居・交易対象物 → 保有・維持・管理 → 増大 対象有蹄類 ungulata 偶蹄目
ウシ科: ウシ・ヒツジ・ヤギ・ヤク
バクトリアラクダ(2瘤) = モンゴル・中央アジア ウマ科: ウマ・ロバ・ラバ (ロバ・ラバは農耕生活利用が一般的) 南米: リャマ・アルパカステップ: 北アジア、中央アジア、カザフ草原、南ロシア草原、パンノニア草原
モンゴル民族
西アジアはステップよりもイラン高原(高地)(1000-1200 m)と呼ぶ方が相応しい
牧童: 12-13歳程度で始める - 能力は外的要因にも関係する 砂漠とオアシスアラビア: ネクト、ルブアルハリアフリカ: サハラ アラブ民族: ◎ラクダ, ヒツジ, ヤギ, ウマ, アラブ馬(天馬・汗血馬・大宛, サラブレッドの原種) 全体として家畜使用目的は、例外多いが、荷役用・乗馬用ではなく乳用である サバンナ: 東アフリカ: タンザニア、ケニア: 多部族 - ウシ ツンドラ/タイガ (tundra and taiga)シベリアSiberia民族 → 北極圏: 古アジア(パレオアジアpaleoasia)語族アジアエスキモー: チェコトカ半島-ベーリング海 (1500人, 1979) 伝統的生業: 海獣猟と毛皮獣猟 言語: エスキモー・アリュート語群 チュクチャ: チェコトカ半島 (14000人, 1979) 伝統的生業: トナカイ飼育、海獣猟、狩猟、漁撈 言語: パレオアジア諸語チェコトカ・カムチャトカ語群 ケレク: ベーリング海沿岸 (100人) → チュクチャやコリャークに同化 言語: パレオアジア諸語チェコトカ・カムチャトカ語群 コリャク(コリヤク, コカギール) カムチャトカ半島 (7900人, 1979) 伝統的生業: トナカイ飼育、漁撈、海獣猟、毛皮猟 言語: パレオアジア諸語チェコトカ・カムチャトカ語群 イテリメン(旧称 カムチャダール): カムチャトカ(先住民) (1400人, 1979) 伝統的生業: 漁猟 言語: パレオアジア諸語チェコトカ・カムチャトカ語群 ニブフ (旧称 ギリヤーク): アムール川下流-サハリン (4400人, 1979) 伝統的生業: 漁猟、海獣猟、狩猟、犬飼育 言語: パレオアジア諸語(独立語) サハ (旧称 ヤクート): サハ共和国および周辺 (456000人, 2002) 北部: トナカイ飼育(半遊牧生活) 南部: トナカイ + ウマ・ウシ飼育 言語: テュルク諸語 エベンキ (ツングース): オビ川中流-オホーツク海沿岸-サハリン(一部、中国・モンゴル) (27000人, 1979)
オロチョン: エベンキの中でアムール川上流に住む一部のグループ エベンキに近い |
伝統的生業: トナカイ飼育、狩猟 言語: ツングース・満州語群 ナナイ (旧称 ゴリド): アムール川・ウスリー川(一部 中国スンガリ川) (10000人, 1979) 伝統的生業: 狩猟、漁撈 (革命後、農業、牧畜、養蜂加わる) 言語: ツングース・満州語群 オロチ: ハバロフスク地方 (1000人, 1979) 伝統的生業: 狩猟 言語: ツングース・満州語群 ブリャート: バイカル湖周辺 (353000人, 1979)
西ブリャート: ロシア人の影響を受け定住生活 ステップ(含む、山岳ステップ) → 遊牧 山岳(西・東南) → ラクダ・ヤク飼育 タイガ → トナカイ飼育、狩猟 言語: テュルク語群 ショル(鍛治タタール): トミ川中流とその支流域 (16000人, 1979) 伝統的生業: 狩猟、漁撈(革命前) + 製鉄・鍛治 言語: アルタイ語族テュルク語群 ヌガナサン: タイムル半島 (900人, 1979) 伝統的生業: 漁撈、狩猟、トナカイ飼育 言語: ウラル語族サモエド語派 ケト: エニセイ川中流-下流 (1100人, 1979) 伝統的生業: 狩猟、漁撈 言語: 親族関係言語なく独立語 ハンティ (旧称 オスチャーク): オビ川中流-下流 (21000人, 1979) 伝統的生業: 狩猟、漁撈、トナカイ飼育 言語: ウラル語俗フィン・ウゴール語派 マンシ (旧称 ボグール): オビ川西側流域-ウラル山脈東側斜面 (7600人, 1979) 伝統的生業: 漁撈、毛皮獣猟 (一部、トナカイ飼育) 言語: ウラル語俗フィン・ウゴール語派 サーミ(ラップ)人: スカンジナビア半島北部ラップランド-ロシア北部コラ半島 ウシ・トナカイ 五畜羊・山羊・牛・馬・駱駝
放牧の要領家族単位 = 家族/家
私有財産所有/消費生活の最小単位
複数家族から成り立つ不定住集団
__4月__→____→____→________→___11月
4月 新芽の出はじめ 移動開始 部族 = 氏族が複数集合したもの |
遊牧文化の歴史(東洋中心)アファナジェーバ文化 Afanasievo culture: エニセイ川上流-オビ川上流 ≈BC 2100- ≈BC1400: 狩猟 + 農耕 + 牧畜 ∵ [ウシ・ウマ・ヒツジ多量発掘(家畜化後形態に近い → 家畜] + [臼発見 → 小麦栽培] + [弓発見 → 狩猟]
+ アラル海特産貝発掘(= アラル海と交流) BC4000末 (BC 3200)-BC 2000初 (BC1900-BC1800) ≈ BC3000: 境に前期後期分割
前期: 狩猟中心。農耕導入 → 後期: 牧畜中心(羊・牛) BC5000-BC4000: 農耕文化 (+ 牧畜: オリエント地帯に始まる) オクニェフ文化: エニセイ川上流 BC2000前半: 青銅器発見。青銅製造術。牧畜初期 アファナシェーバ文化の継承 アンドロノボ文化 Andronovo culture: 中央アジア草原地帯(オビ川-エニセイ川, カスピ-アム・シル + ウラル川・ボルガ川・ドン川) BC2000前-BC1000初 - ケルチェミナール文化の継承 前期: 第1期 = BC 18c-16c + 第2期 = 15-13
ウシ・ウマ(荷役用か騎馬用)・ヒツジ・ヤギ・ブタ +
ヒツジ・ヤギ増加(多) (小型) + 馬具(轡、青銅器)発見 (乗馬目的) カラスク文化 Karasuk culture: エニセイ川-オビ川上流 BC13c-BC8c: アンドロノボ文化後期に重なる ヒツジ(多量)・ウマ・ウシ・ラクダ 馬は後期に馬具が発見され、更に石画に幌付四輪車の絵が確認され、乗馬用とし利用したと考えられる。幌付四輪車から移動生活も推定される 既発見の青銅器を元に影響源を調べているが確定していない 集落跡・灌漑跡-農耕生活 カラスク文化の人々は、牧畜人、農耕人の2タイプがあったと予測される 木槨墳文化 Timber-grave culture: 南ロシア草原(黒海-カスピ海) BC1900-BC1200: クルガン(古墳/高塚/高塚墳, kurgan) = スルプナ文化 Srubnaya culture アンドロノボ文化と木槨墳文化は酷似 - 交流存在? Ex. 短剣・刀子・斧木槨墳文化成立の歴史
B.C.3000年紀末: 堅穴墳文化 - 狩猟・漁労
BC3000年紀末-
スキタイ文化 Skythai culture: ギリシア記録からは野蛮と指摘BC2000年紀前半: 横穴墳文化 - 牧畜・農耕 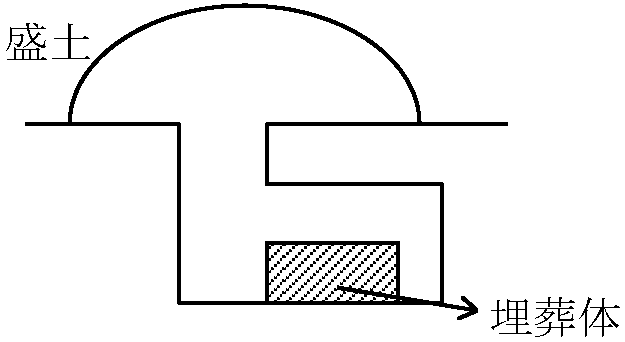 馬具未出土 → 牧主農副 BC8c-BC1c 全盛 ⇒ アルタイ: 鉄(製鉄炉)・金細工出土 BC7c-BC3c 木槨墳文化を継承するもの
スキタイ人 ヒツジ・ヤギ・ウシ・ウマ(馬具確認) 墓(クルガン): 大規模なもの発見。馬・殉葬。モンゴロイド BC5C-BC3C スキートシベリア文化 農耕・灌漑・集落跡(既にスキタイ文化で確認された) + 騎馬も確認 BC7C-BC4C マイエミール文化 Mayemir: タガール文化と殆ど同じ特徴 オビ川上流アルタイ地方
BC3C-BC2C パジリク文化 Pazyryk culture
オビ川上流(現モンゴル・ソ連国境の西側)
BC3C? パジリク古墳群 Pazyryk burials
大型円墳5基含む8基の墳墓発掘調査 モンゴル地方の遊牧文化 (未詳)綏遠文化 Suiyuan culture:BC5c-BC2c 内蒙古綏遠地方 ≈ エニセイ・タガール・オビ・バジリク地方文化
∵ 青銅器・鉄器等類似 匈奴の冒頓単干は幼少期に人質として月氏に囚われていた 月氏脱出後に父を殺し単干となる(BC 209) 当時、近隣に勢力をはっていた東胡を滅ぼし征服 タシュトゥィク文化 Tashtyk culture: エニセイ地方。タガール文化に続くもの BC1c-AD5c モンゴロイド(ユーロペオイド) - モンゴル勢力の西方侵入 遊牧(ヒツジ・ヤギ・ウシ・ウマ) = 遊牧民自身
↔ 農耕 (キビ) = 捕虜となった農耕民 タガール文化後継 ↔
綏遠文化(匈奴文化)青銅器・鉄器発見 - 漢・匈奴文化とすべき?
オンドル(ペチカに似た暖房具)を持つ家発見 |
BC 99 李陵の匈奴遠征(失敗) 李陵捕らえられる → 陵 = 李陵 (説) 板石墓(立石墓)文化: ザバイカル地方BC 7c-BC 3c: 青銅器・鉄(製)器。丁霊文化と推定される ノインワラ文化 BC 1c-AC 1c 匈奴文化 丁霊 / 高車丁霊 6c 突厥(トルコ系): 回鶻→セルジュクトルコ→オスマントルコ→トルコ共和国スキタイの歴史スキタイ以前の先住民キンメリア人(アッシリア、サルゴンII世(BC 722-705)の資料に見られる)BC 714 ウラルト城塞都市攻撃 BC 680-BC 670頃: スキタイ圧迫避けコーカサス北部から南部Origentへ移動
スキタイに追放された後、Orient, Turkeyへ移動。スキタイもキンメリア人を追いアッシリアへ侵入 ソビエトの研究: キンメリア人もイラン系、スキタイと共に遊牧 元来、スキタイ・キンメリアは同一の民族であったものが部族連合体をなす スキタイの進出ヘロトドス「歴史」にスキタイ登場。「歴史」全9巻中第4巻に詳述BC 514: アケメネス朝ペルシア、ダリウスI世がスキタイ遠征失敗(ヘロトドスは他に「ペルシア戦争史」記す) BC 4cまで繁栄 (北海北岸原住) BC.7c: ポルガ西方進出, キンメリア人追放 BC 6c: 定住 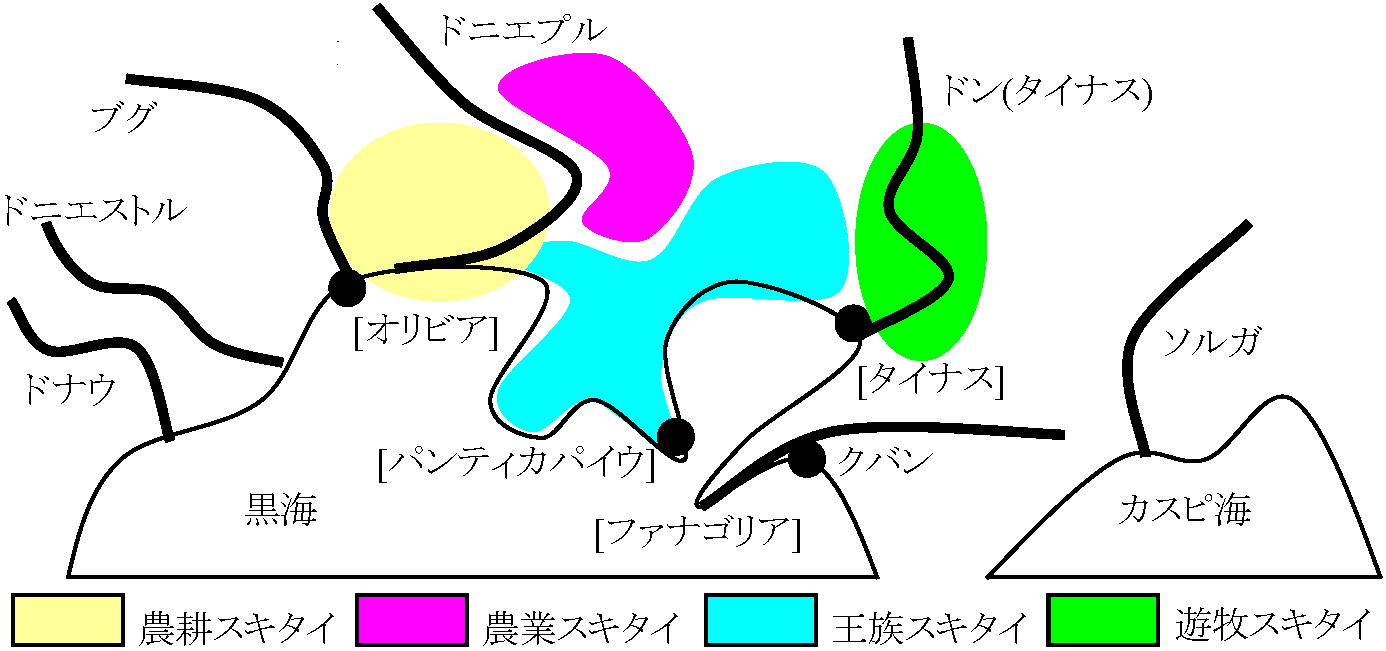 「歴史」によると ⇔ 黒海北岸スキュティア地方にスキタイは住む タナイス(ドン)川を超えるとサウロマタイ(サルマート)とされる人々の土地となり、スキタイは住まない 黒海北海にはギリシア植民地がかなり有り、そこと交易が記され、その頃スキタイはスキタイ王国と名乗った スキタイ発展Pax Scythica = 「スキタイの平和」 - スキタイ発展をギリシアで象徴した言葉ギリシア植民地との交易
スキタイ_________⇔___ギリシア殖民都市 スキタイ衰退2-3 c以後未詳となる
メディア地方: BC 7-BC 3c (7-7c少、5-4c中心)
ドニエプル川下流
キロボクラード
BC 7-6c 文化遊牧: 死者と一所にクルガンにはしばしば馬が埋葬される
クバン川上流マイユープのウイスキークルガン(BC 6c)で360頭の馬発見 - 馬具も発見される トルコ系諸民族の拡散ウイグルギルギス ハザール マジャール ペチェネーグ カラハン朝 Kara-Khanid Khanate カズナ(カズニ)朝 Ghaznavid dynasty セルジュク オスマン モンゴル系諸民族の拡大契丹と遼契丹人、耶律氏遼(916-1125) 契丹(647以前, 983-1066): この間の国号, これ以外は遼 西遼(1132-1218)現トルキスタン - 金に滅ぼされるタタール (韃靼, Tatar): テュルク系民族(トルコ系諸民族) モゴール Moghol: アフガニスタン、ヘラート州定住モンゴル系民族 近世・近代のモンゴル系・トルコ系諸民族オイラート Oiradジュンガル カザフ キルギス ウズベク モンゴル革命 (1921) |

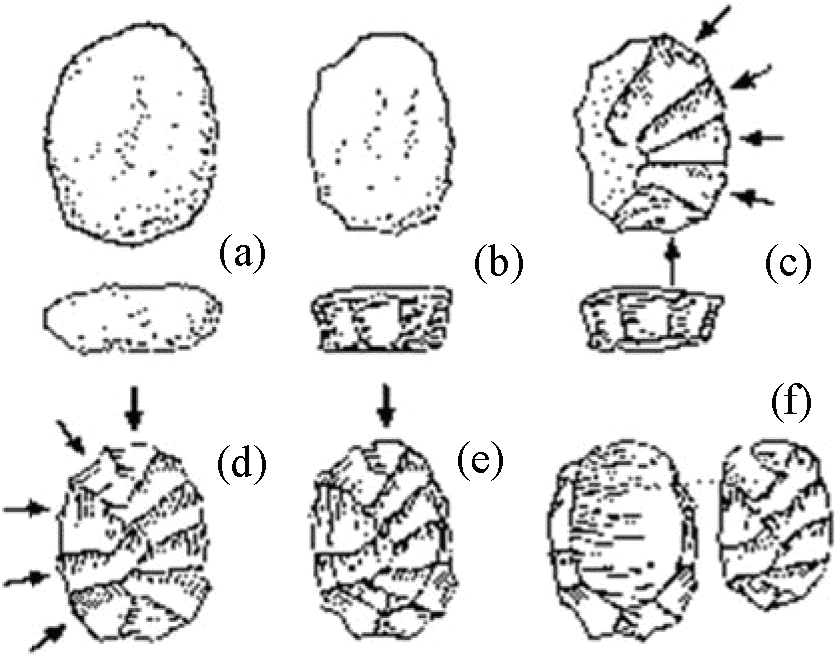 ルボロア技法による石器: 前期旧石器時代の技法
ルボロア技法による石器: 前期旧石器時代の技法