(Upload on Marach 3 2026) [ 日本語 | English ]
Mount Usu / Sarobetsu post-mined peatland
From left: Crater basin in 1986 and 2006. Cottongrass / Daylily
HOME > Lecture catalog / Research summary > Glossary > Geological timetable > Fossil
Fossilization (化石化)the process of becoming a fossilOrganism dies ● ↙ ↓ ↘ Covered Decomposed Destroyed (sediment, ice, amber, etc.) (biological factors) (physical factors) ↓ ↘ ● Destroyed Fossilized ↘ (physical and ↓ ↘ ↘ biological factors) Destroyed Stays buried Discovered (moved, (not exposed, (exposed melted, eroded not found) and found) or crushed) ● Fig. Diagram showing the process of fossilization. Conditions necessary for fossilization
|
Methods of fossilization
|
|
Def. the study of fossil fungi Phanerozoic (顕生代): a great diversity of fossil fungi |
|
In the 1950's laboratory experiments showed that electrical discharges, like lightning, may have caused chemical compounds on the early earth to combine, forming the building blocks for life. Since then, theories for how life began have explored numerous paths. Early in the earth’s evolution the chemicals necessary for life existed in the oceans or atmosphere, but for hundreds of millions of years the Earth was probably lifeless. If lie had evolved, bombardment by asteroids and comets would have vaporized the oceans, sterilizing the planet. Asteroids and comets likely brought organic compounds with them. Heavy bombardment probably ended 3.8 billion years ago. Rocks about this age contain organic carbon, suggesting life arose quickly. Rocks 3.5 billion years old contain the first fossils of simple (prokaryotic) cells that lack a nucleus. These cells were the only life forms until about 2 billion years ago when (eukaryotic) cells with a nucleus appeared. |
Stromatolite (ストロマトライト) |
[ origin of life ]
PrecambrianChemical fossil (化学化石): any of various organic compounds found in ancient geological strata ta that appear to be biological in origin and are assumed to indicate that life existed when the rocks were formed.Archaean strata containing chemical fossils - life existed over 3500 m.y.a, perhaps even as much as 3800 Mya 3100 Mya: degradative products from chlorophyll (Oro et al. 1967)2000 Mya: fossils of amino acids 1967 Barghoorn: microbe fossils
Collected from Fig Tree Formation in southern Africa similar with Cyanophyceae |
Eukaryotes (真核生物)Proterozoic Eon (21 billion years ago, bya) Grypania - tube-shaped fossil
(size > 1 cm) ≠ prokaryotes Symbiotic theory (共生説)eukaryotes were evolved by symbiosis - supported by fossil recordsEdiacara (or Ediacaran) fauna (エディアカラ動物群)= Ediacaran biota (formerly Vendian), 635–538.8 myacharacterized by enigmatic tubular and frond-shaped, mostly sessile, organisms 
|
Aquatic organismsall the fossils of organisms were aquatic or oceanic before SilurianDiatomsfossil records: appeared after Jurassic and insreased in Cretaceous, as well as Dinoflagellates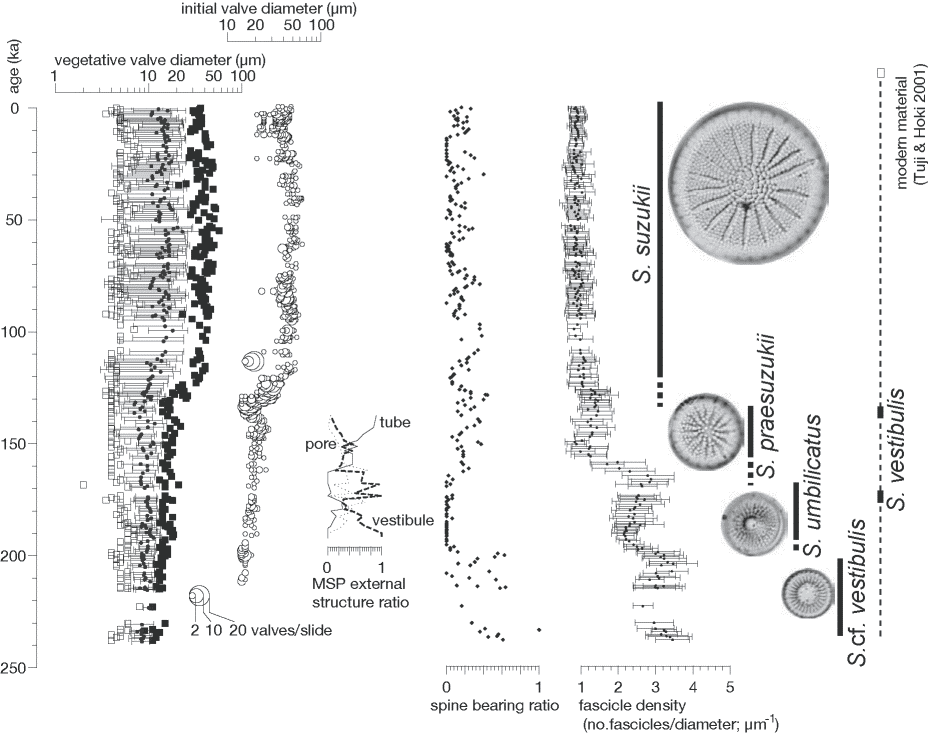 Fig. 4. Stratigraphic changes in valve morphology of Stephanodiscus species, which occurred monospecifically. From left to right; valve diameter, open squares (minimum), solid small circles (average) and solid squares (maximum); distribution of initial valve diameter; frequency of vestibule, pore or short tube for mantle strutted process external opening, vestibule in dashed line, pore in dotted line and tube in solid line; spine-bearing valve ratio, broken line showing the exceptionally high values intercalated by S. vestibulis's occurrence; distribution of fascicle/valve diameter ratio called fascicle density in this study; schematic diagram showing biostratigraphic ranges and phylogenic hypothesis for Stephanodiscus cf. vestibulis, S. umbilicatus, S. praesuzukii and S. suzukii in Lake Biwa with reference to S. vestibulis. (Saito-Kato et al. 2015) |
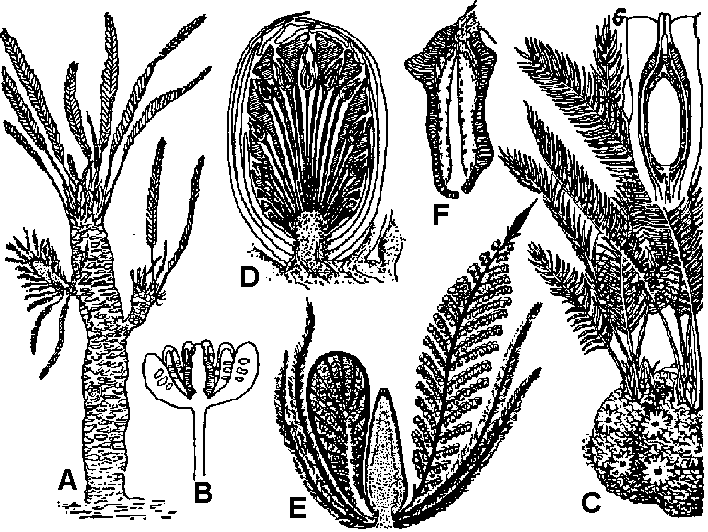
Green algae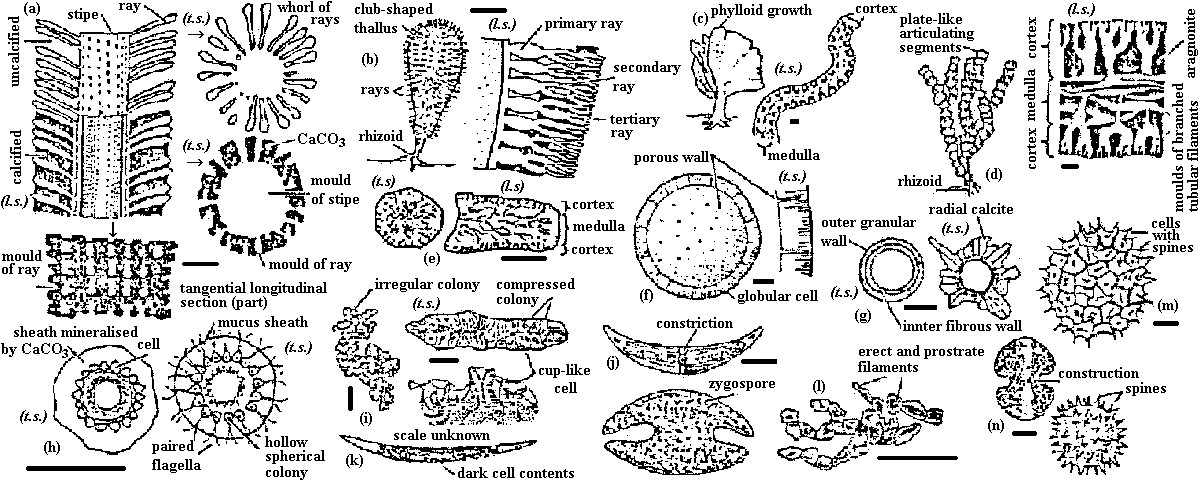 Fig. (a-e) Calcareous green algae. (a) Oligoporella, (b) Petrascula, (c) Eugonophyllum, (d) Halimeda, (e) Boueina. l.s. and t.s. = longitudinal and transverse sections. (f-n) Green algae. (f) Tasmanites, (g) Calcisphaera, left = non-radiosphaerid, right = radiosphaerid, (h) Eovolvox, left, as preserved in CaCO3, right, reconstruction with inner daughter colony, (i) Botryococcus, left = colony, right = sections through tallus, (j) Recent Closterium (above), with zygospore, (below), (k) fossil Closterium-like desmid (scale unknown), (l) endolithic chaetophoralean-like alga, (m) Pediastrum, (n) recent Cosmarium with zygospore to right. 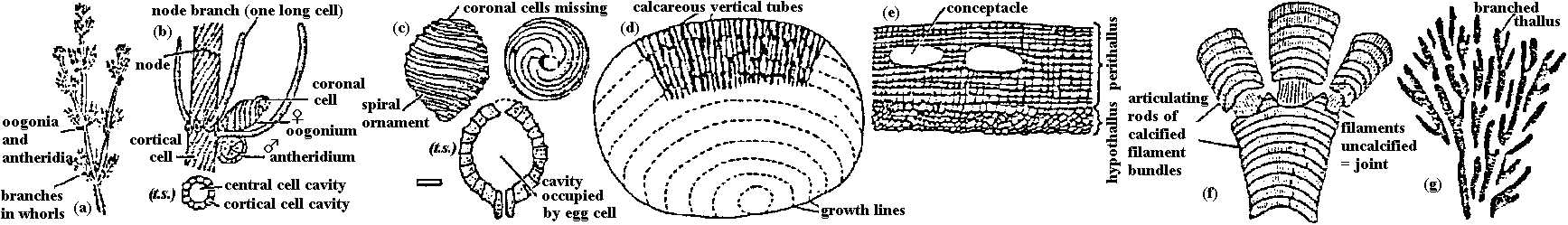 Fig. a-c: Charophytes. a: Chara plant, b: Detail of stem, branches, oogonia and antheridia, c: Stellatochara progonite (bar = 100 μm). d-g: Calcareous red algae. d: Solenopora, e: Lithothamnion, f: Corallina, d: Epiphyton. Phaeophyceae (褐藻): appeared after Trias Rhodophyceae (紅藻): flourished in Ordovician Alteration of generationAll vascular plants: haploid generation = gametophyte, diploid generation = sporophyte- life cycle = biphassic system Life cycle of Ulva: G = Sp. → primitive Most plant species: G << Sp
ultrastructure: antrophic, heterotrophic |
☛ Epoch: Paleozoic
Devonian (デボン紀, 4.16-3.67 Bya)Carboniferous (石炭紀, 367-289 Mya)Equisetidae, Equisetopsida (s.s.) or Sphenopsida トクサ類GinkgoG. biloba: Permian (270 mya) - present (living fossil)
by Jurassic: worldwide |
Fossil species: G. adiantoides, G. apodes, G. cranei, G. digitata, G. dissecta, G. gardneri, G. ginkgoidea, G. henanensis, G. huolinhensis, G. huttonii, G. yimaensisPermian (二畳紀, 290-251 Mya) |
Rhynie flora (ライニーフロラ)early DevonianRhynia major, R. gwynne-vaughani, Horneophyton lignieri, Asteroxylon mackiei, etc.  Fig. A reconstructed example of landscape in the Devonian Rhynie chert Four major floral zones in the PaleozoicEuramerican flora (ユーラメリカフロラ)≈ 3 Bya (late Paleozoic): present Europe and North AmericaLepidophytina: Lepidodendron (鱗木),Sigillaria (封印木) Articulatae: Calamites (蘆木) Fern-like foliage: Neuropteris, Alethopteris, Taeniopteris, Callipteris, Gigantopteris Conifer: CordaitesAngara or Siberia flora (アンガラフロラ)Cathaysia flora (カタイシアフロラ)
Gigantopteris: characteristic distribution after the Permian Gondwana flora (ゴンドワナフロラ)recorded from glaciers developed in the upper Permian |
Glossopteris: leaf = main vein (costa), lateral vein – dichotomous branching 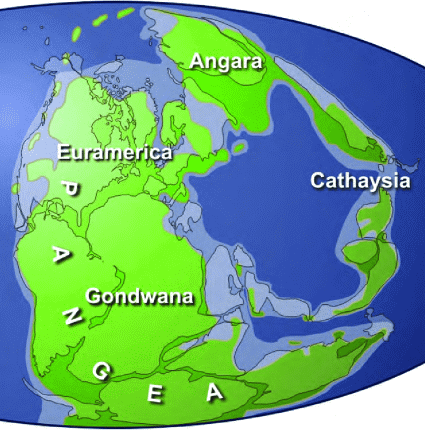 Fig. 1. The Late Paleozoic supercontinent, Pangea. Four major floral zones are indicated, tropical Euramerica and Cathaysia, and temperate Angara and Gondwana (Scotese 1997)
Fig. 1. The Late Paleozoic supercontinent, Pangea. Four major floral zones are indicated, tropical Euramerica and Cathaysia, and temperate Angara and Gondwana (Scotese 1997)
|
Seed fern

Alethopteris Sternberg
Pennsylvanian, Minto Formation
Rothwell, New Brunswick
Collector: WB Evans
Seed fern

Alethopteris Sternberg
Pennsylvanian, St. Clair,
Pennsylvania
United States of America
Collector: WH Forbes
Leaves of Calamites

Annularia Sternberg
Pennsylvanian, Clifton Formation
Clifton, New Brunswick
Collectors: RF Miller and
J McGovern, 1994
Giant horsetail plant

Calamites Suckow
Pennsylvanian, Tynemouth
Creek Formation
Gardner Creek, New Brunswick
Collector: F Sherwood, 1989
Plant

Psilophyton charientos Gensel
Devonian, Campbellton Formation
Dalhousie Junction, New Brunswick
Collector: PG Gensel, ca 1986
Plant

Lepidodendropsis Sternberg
Mississippian, Albert Formation
Bloomfield, New Brunswick
Collector: RF Miller
(St John Museum, Sept 19 2014)
The floras chronologically differed between the Northern and Southern Hemisphere.
The geological distribution of Carboniferous and Permian plants.
| Devonian | Carbon | Permian | Triassic | |||||
| Lower | Upper | Low | Upper | |||||
| Westphalian | Stephanian | |||||||
| Spehnopsida | ||||||||
|---|---|---|---|---|---|---|---|---|
| Asterocalamites | ===§=== | ==== | ||||||
| Calamites | ======= | ====§==== | ======== | ======= | === | |||
| Asterophyllites | ======== | ======== | ======= | |||||
| Annularia | ======== | ======== | ======= | |||||
| Lobatannularia | ===== | ======= | ----------- | ----------- | -> | |||
| Schizoneura | ===== | ======= | ======= | === | ||||
| Phylletheca | ==== | ======== | ======== | ======= | ======= | ======= | ==> | |
| Euisetites | ======== | ======== | ======= | ======= | ======= | ==> | ||
| Sphenophyllum | ======= | ======= | ======== | ======== | ======= | ===> | ||
| Cheirostrobus | ======= | |||||||
| Lycopsida | ||||||||
| Lepidodendron | ===== | ======= | ====§==== | ======== | ======= | |||
| Lepidopholois | ===== | ====§==== | ||||||
| Sigillaria | ==== | ====§==== | ======== | == | ||||
| Stigamaria | ===== | ======= | ======== | ======== | ======= | ==-- | ||
| Archaeosigillaria | ===== | ======= | ||||||
| Bothrodendron | ===== | ====§=== | = | |||||
| Aselanus | ======== | ------ | ||||||
| Lepidocarpon | ======= | ======== | ||||||
| Miadesmia | ======== | |||||||
| Lycopodites | ======= | ======== | ======== | ======= | ======= | ======= | ==> | |
| Selaginellites | ===== | ======== | ======= | ======= | ======= | ==> | ||
| Pteropsida | ||||||||
| Coenopterideae | ||||||||
| Botryopteris | ======= | ======== | ======== | ======= | ||||
| Metaclepsydropsis | ======= | ======== | ======== | = | ||||
| Stauropteris | ======= | |||||||
| Diplolabis | ======= | ======== | ||||||
| Botrychioxylon | ======= | |||||||
| Etapteris | ======== | |||||||
| Clepsydropsis | ======= | ======== | ======== | ======= | ||||
| Ankyropteris | ======= | |||||||
| Asterochlaena | ======== | ======== | ======= | |||||
| Other ferns | ||||||||
| Thamnopteris | ===== | ======= | ||||||
| Psaronius | ======== | ======== | ===§=== | |||||
| Rhacopteris (Anisopteris) | ===§=== | |||||||
| Oligocarpia | ====§==== | ======== | ======= | |||||
| Spermatophyta | ||||||||
| Pteridospemales | ||||||||
| Pecopteris | ======== | ====§==== | ======= | ======= | ======= | |||
| Cladophlebis | ======== | ======= | ======= | ======= | ==> | |||
| Alethopteris | ====§==== | ======== | ======= | |||||
| Lonchopteris | ====§==== | ======== | ----------- | ----------- | ------------ | |||
| Callipteris | ======= | ======= | ||||||
| Odontopteris | ==== | ======== | ===§=== | |||||
| Callipteridium | ======== | ===§=== | ||||||
| Neuropteris | ==== | ====§==== | ||||||
| Neuropteridium | ======= | ======= | ======= | |||||
| Gondowanidium | ==== | ======= | ||||||
| Linopteris | ====§==== | ======== | ======= | |||||
| Adiantites | ======= | ===§=== | ======== | --------------- | ----------- | ----------- | ------------ | --> |
| Sphenopteris | ======= | ======= | ======== | ======== | ======= | ======= | ======= | ==> |
| Mariopteris | ====§==== | == | ||||||
| Sphenopteridium | ===== | ======= | ||||||
| Rhodea | ===== | ===§=== | ======== | |||||
| Cardiopteris | ======= | |||||||
| Gigantopteris | ===== | ======= | ||||||
| Taeniopteris | ====== | ===§=== | ||||||
| Thinnfeldia | ======= | ======= | ======= | ==> | ||||
| Gangamopteris | ===== | ======= | ||||||
| Glossopteris | ===== | ======= | ======= | ======= | ||||
| Chiropteris | ======= | ======= | ======= | ==> | ||||
| Stems, etc. | ||||||||
| Lyginopteris | ======= | ======== | ||||||
| Heterangium | ======= | ======== | ======== | |||||
| Rhetinangium | ======= | |||||||
| Megaloxylon | ======== | |||||||
| Calamopitys | ======= | |||||||
| Stenomyelon | ======= | |||||||
| Protopitys | ======= | |||||||
| Cladoxylon | === | ======= | ||||||
| Medullosa | ======== | ======== | ======= | |||||
| Cycadales & Bennettitales | ||||||||
| Dioonites | ======= | ======= | ======= | ==> | ||||
| Pterophyllum | ==== | ======== | ======= | ======= | ======= | ==> | ||
| Sphenozamites | ======= | ======= | ======= | ==> | ||||
| Ginkgoales | ||||||||
| Baiera | ==== | ======= | ======= | ======= | ==> | |||
| Sanortaea | ==== | ======= | ||||||
| Cordaitales | ||||||||
| Cordaites | ======= | ======== | ======== | ======= | ------------ | ------------ | --> | |
| Pitys: Archaeopitys | ======= | |||||||
| Coenoxylon | === | ===== | ||||||
| Dolerophyllum | ======== | ======== | ======= | |||||
| Coniferales | ||||||||
| Araucarites | ===§=== | ======= | ======= | == | ||||
| Walchia | ==== | ======== | ======= | |||||
| Ernestia | ======= | |||||||
| Voltia | === | ======= | ======= | |||||
| Pityanthus | ======== | |||||||
| Pityospermum | ======= | ======= | ||||||
| Dicranophyllum | === | ======== | ======== | ===§=== | ======= | |||
| Ullmannia | === | ======= | ||||||
| Gemphostrobus | ======= | === | ||||||
| Plants of uncertan position | ||||||||
| Psygmophyllum | === | ======= | ======== | ======== | ======= | === | ||
| Plagiozamites | ======== | ======= | ||||||
| Tingia | ==== | ======= | ||||||
| Pelourdea | ==== | ======= | ======= | ======= |
|
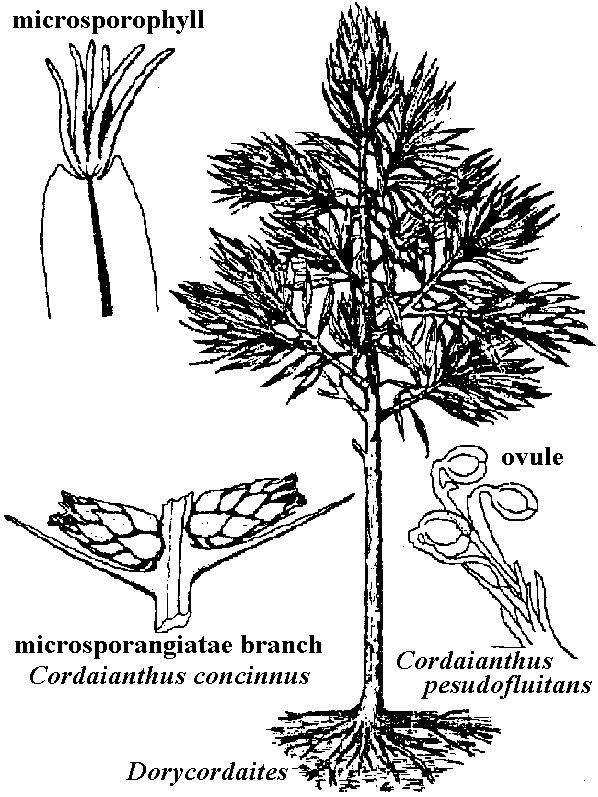
Cordaites trees are an extinct group of seed-bearing conifer-like plants. Although an important part of the Pennsylvanian landscape, they only made up ten percent of the forest biomass. Cordaites trees, up to 30 meters tall, grew on the lowlands, while shrub varieties and shorter trees with stilt-like roots lived in swamps along the edges of seashores and estuaries. Like many fossil plants, different parts of the tree may are known by different names. The former genus “Dadoxylon” refers to fossilized wood and includes logs that likely belonged to cordaites trees. The strap-like leaves, known as Cordaites, vary in size from a few centimeters to one meter in length. The unfertilized seeds, Cardiocarpus, were borne on the leafy branches of the tree and are often found in large numbers. (St John Museum, Sept 19 2014) |
|
Macro-fossil (巨大化石)Taxonomical keytracheid (仮導管)cuticle (角皮) + stoma (気孔) spore formation (胞子形状) branching pattern (軸分枝系) sexual tissues (繁殖組織): Gymnospermae Ex. cone = pine Taxonomical classificationDivision Rhyniophyta (ライニア)= Division Pteridophyta, s.l.Found in the Early Devonian (≈ 419-393 mya) Subdivision Psilophytina (裸茎植物)Class Rhyniopsida (ライニア)= Class Psilophytopsida (古生マツバラン)1970 Banks: three major groups =
Rhyniales + Order Rhyniales (ライニア)stele = centrarch 心原型 (≈ endarch 内原型)lacking secondary vascular bundle tissues sporangium - split vertically Rhyniaceae (ライニア) Rhynia Kidst. et Lang, Horneophyton Bargh. et Darrah (= Hornea Kidst. et Lang) 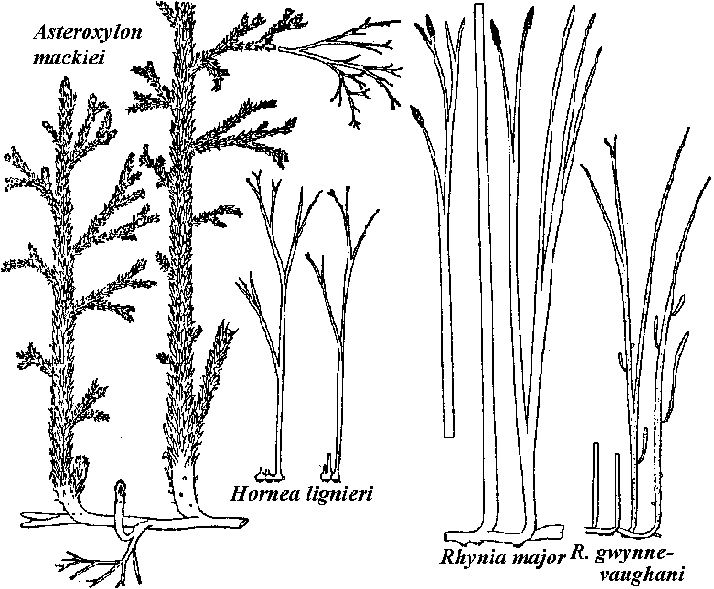 Rhynia: R. major (2n) → same species (?) → (n) R. gwyene-vaughan 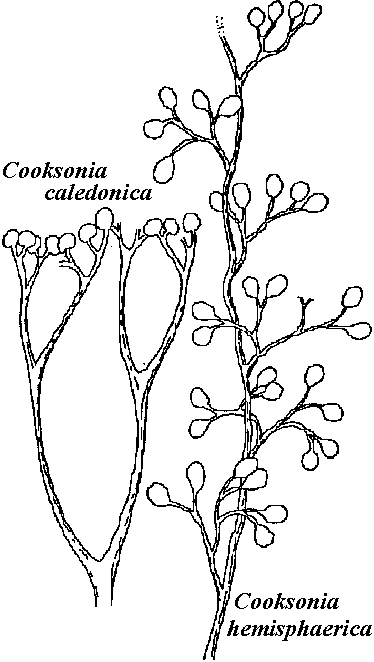
Hypothesis: R. gwyene-vaughan: developing inconstant branch (不定枝) → R. major Dawasonites Halle Trimerophyton Hopping (トリメロフィトン) three branching → evolving to Equisetum Cooksonia Lang Asteroxylaceae (アステロキシロン) Protolepidodendron Krejči (Gr. protos = first, lepidos = scale, dendron = tree) 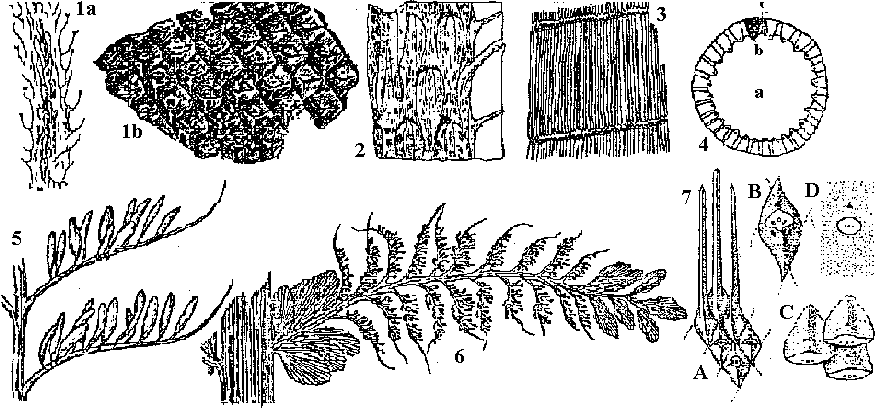 Fig. 1.Protoledpidoendron scharyanum (Mid-Devonian, Germany. Lower Devonian, Yungnagn, China) a: branch attached with thread-like foliage, b: magnified branch surface. 2.Lepidodendron oculis-felis (Lower Permian). 3. Calamites cisti (Upper Permian, Germany). 4. Calamites, a: medullary groove (髄孔), b: xylem, c: medullary ray (射出髄). 5. Archaeopteris latifolia. 6. Archaeopteris hibernica. 7. Leaf based of several Lycopods: a, young twig of arborescent Lycopod; b, Lepidodendron; c, Lepidophloios; d, Bothrodendron Calamophyton: mid-Devonian → morphologically similar with Hyenia = classified into Arthrophyta, homospore or heterospore is unknownSphenophyllum (Gr. sphenos = wedge, phyllum = leaf): tree fern (Articulatae, 有節類). strobilus = Bowmanites. homospore Order Zosterophyllales (ゾステロフィルム目)lateral sporangia (側生胞子嚢) → sporangium = transverse dehiscence
|
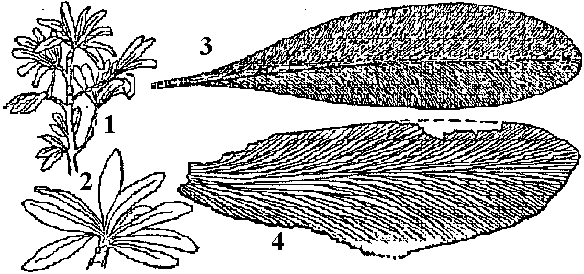
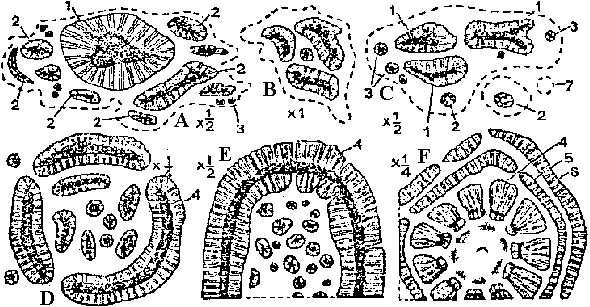
[the present ferns] ZosterophyllaceaeBucheria Dorf, Zosterophyllum Daws. 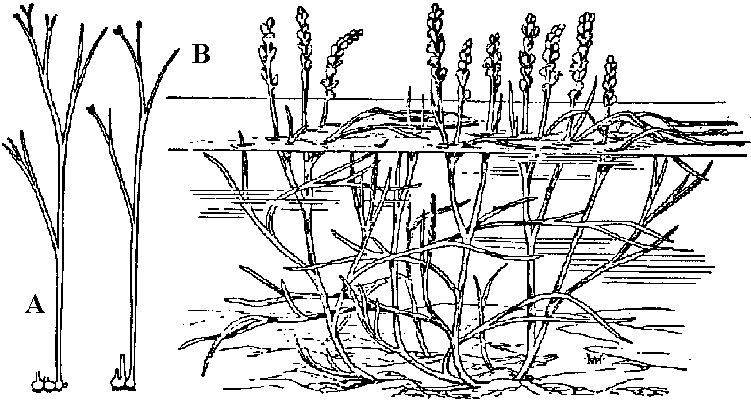 Fig. A: Horneophyton lignieri. B: Zosterophyllum rhenanum Kräusel u. Weyland. Upper Devonian (reconstruction) (Weyland 1935) Zosterophyllum: aquatic plant (mesic sites), not developing leaves 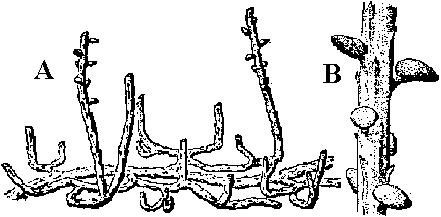 Upper Devonian. Fig. Kaulangiophyton akantha. A. Restoration showing creeping axes with erect branches, two of them fertile. B. Restoration of part of fertile axis. (Gensel, Kasper & Andrews 1975). Upper Devonian. Fig. Kaulangiophyton akantha. A. Restoration showing creeping axes with erect branches, two of them fertile. B. Restoration of part of fertile axis. (Gensel, Kasper & Andrews 1975).[Division Rhyniophyta, described above] Division Pteridophyta (シダ植物)Subdivision Psilophytina (裸茎植物)Class Psilophytopsida (古生マツバラン)phylogenetic relations are unknown (taxonomical group of convenience)Order Psilophytales (プシロフィトン/古生マツバラン)Silurian - PermianPsilophytaceae (プシロフィトン): Psilophyton Daws. Fossil records from chert: Rhynia, Psilophyton, Taeniocrada, Cooksonia, Sporogonites Body plan (structure, 体制): developed vasular bundles, not differantiated to stems and leavs Subdivision Lepidophytina (小葉植物)= Microphyllophytina (小葉植物), Lycopodiinae, Lycophytina, LycopsidaClass Aglossposida (無舌)Order Protolepidodendrales (古生ヒカゲノカズラ)early Devonian - ending Devonian (partly until Carboniferous)maostly herbs = secondary growth unproved Drepanophycaceae: Baragwanathia, DrepanophycusProtolepidodendraceae: Arachaeosigillaria Kidston, Lepidodendropsis Lutz, Protolepidodendron Krejči Lycopodiaceae (2 species in existence): after Carboniferous Fossil species: Lycopodites Brongn, Paurodendron Fry Class Glossopsida (有舌)Subclass Primofilicidae* (原シダ)Order Protopteridales* (古生シダ): Devonian-Carboniferous Protoptericaceae* (古生シダ): Protopteridium, SvalbardiaOrder Cladoxylales* (クラドキシロン): mid-Devonian-Carboniferous only sporophyts reported, erect above-ground stem, dichotomous or irregular branching, crown developed on the top Fern-like foliage (羊歯的葉)Taxonomical and phylogenetical positions are not clearly determined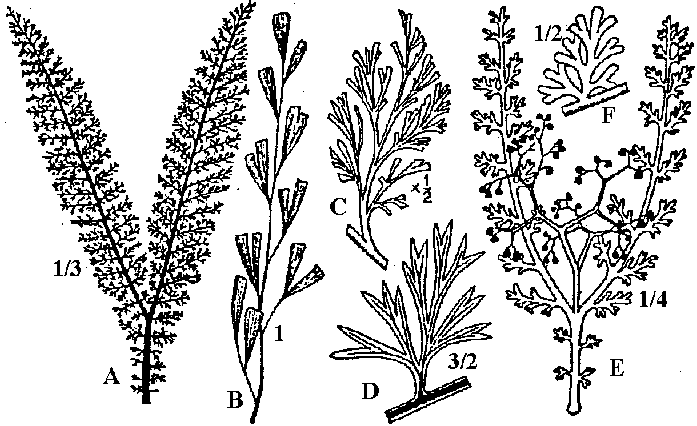 Fig. Lower Carboniferous pteridosperm fronds (石炭紀下部シダ状種子植物). A, Sphenopteridium capillare, complete frond. B, Adiantites machanekii, fragment of frond showing form of ultimate segments. C, Sphenopteris affinis, portion of frond. D, Rhodea smithii, ultimate pinna. E, Diplopteridium teilianum, reconstruction of complete frond with Telangium type of pollen-bearing organs attached. F, pinna of Sphenopteris type, from Diplopteridium teilianum.  Fig. Upper Carboniferous peridosperm fronds. A, Mariopteris, pinna showing double dichotomy. B, Mariopteris pinnule. C, Sphenopteris pinnule. D, Pecopteris (Asterotheca) daubrei, pinnule. E, Pecopteris armasii, pinnule. F, Odontopteris, complete frond. G, Odontopteris pinnule. H, Alethopteris, apex of frond. I, Aletopteris pinnule. J, Lonchopteris pinnule. K, Neuropteris, apex of frond. L, Neuropteris pinnule. M, Linopteris pinnule. |
Telome theory (テローム説)1930 Zimmerman, Walter Max 1892-1980: proposedDef. telome: ultimate terminal portion of a dichotomising axis
fertile telome = terminated by a sporangium ↔ sterile telome |
Processes of developing telomes
i) overtopping. ii) reduction. iii) planation. iv) syngenesis or webbing. v) curvation Origin = Rhynia → divergence (monophyletic theory) |
Heterospory (異型胞子)origin of seed: gymnospermae vs angiospermae ⇒ monophyly or polyphyly?Homosproy and heterosporyFern → hydro-fern = heterospory |
Embryo – leaves keeping spores
macrosporangium → [sporocyst] → megaspore |
Origin of angiosperms (被子植物起源)Cretaceous age: angiosperms appeared suddenly1879 Darwin: abominable mystery (in a letter to Hooker J) 1927 Knowlton, Frank Hall (1860-1926): In Plants of the past from the time of their appearance they did not progress at all due to their full-fledged appearance 1969 Bailey & Takhtajan: missing link as the polyphyletic origin1970 Axelrod: origin was in mild uplants at low latitudes 1970 Smith: origin = southeast Asia near to Malaysia 2003 APG II: supported monophyletic origin(*: fossil record = extinct) Seed plants in mesozoicDivision Spermatophyta 種子植物Subdivision Gymnospermae 裸子植物Class Cycadopsida ソテツSubclass Pteridospermidae* シダ状種子植物Devonian - Jurassic (fossil record)integument (珠皮) formed by telome
= Cycadofilicales ソテツシダ目 (seed ferns): sometimes not established
considered to be the most primitive seed plant reported from fossils
Medullosa Cotta: upper Carboniferous - Permian |
Calamopitys Unger: fissil of stem Peltaspermaceae*: Permian - Triassic Lepidopteris Schimp.: fossil of leaf Peltaspermum Brongn.: fossil of female reproductive organ Corystospermaceae: Triassic Pterorachis Freng.; ♂ reproductive organ Caytoniaceae: Triassic -Cretaceous Caytonia Thomas = macrosporophyll (♀)
= Sagenopteris: leaf Glossopteridaceae: Carboniferous - Permian → distributed mainly in Gondwana land (Order Glossopteridales, when established) Gangamopteris McCoy: leaf fossil Glossopteris Brongn.: leaf fossil Subclass Cycadidae ソテツOrder Bennettitales*: Triassic- end of Mesozoic)
Williamsonia Carr. (Bennetiocarpus: seed fossil, Bennettistemon: male inflorescence) Wielandiellaceae* Wielandiella Nathorst (= Anomozamites Schimper) Williamsoniella Thomas (= Nilssoniopteris Nathorst) Cycadeoideaceae Cycadeoidea Buckland Order Cycadales ソテツNilssoniaceae*: Jurassic - Cretaceous1933 Florin, Rudolf (1894-1965): upper Triassic in Sweden named Bjuvia simplex - most primitive cycas 1961 Harris TM: reconstructed Nilssonia1975 Asama & Sekido: established Nilssoniaceae with Nilssoniocladus
= Nilssoniocladus: long and spur shoots Subclass Pentoxylidae ペントキシロン*Order Pentoxylales*
Class Coniferopsida (球果植物)Order CordaitalesUpper Devonian - Permian, many fossils collected from the Northern Hemispheretall tree branching at the upper part, slim stem, large and clear pith leaf: spiral arrangement, spoon-like, macrophyllous, single leaf ≈ 1 m long Two plausible evolutionary pathways (similar with Araucariaceae)
|
[Epoch: Triassic 2.51-1.95 Bya]
|
Table. Geological distribution of some Triassic and Rhaetic plants
Paleozoic
Trias-
-Rhaetic
|
Order Corystospermales Petriella 1981 (Umkomasiales) Corystospermaceae (Umkomasiaceae) endemic to the Southern Hemisphere, including Inida Dicroidium (seed fern), leaves
Kykloxylon and Rhexoxylon, wood Order Peltaspermales Taylor 1981 mostly in Jurassic Lepidopteris Schimper 1869, leaves Dipteridaceae flora (ヤブレガサウラボシ植物群)middle and late Triassic - early JurassicDistribution of Dipteridaceae: central Europe - North America Class Pinopsida (球果), most of them are form genera Incertae sedis (form genus by leaves) - because of no cones Brachyphyllum: like modern Podocarpaceae and Cryptomeria Pagiophyllum: like modern Araucaria bidwillii Cupressinoxylon: like modern Cryptomeria or Podocarpaceae Ginkgoitaceae: like modern Araucaria bidwillii or Cryptomeria Elatocladus: like modern Cephalotaxus harringtonia or Metasequoia glyptostroboides Cuppressinocladus: like modern Cupressaceae Pityocladus: like modern Pinaceae (only leaf trace = Pityophyllum) Podozamites: like modern Nageia nagi, Agathis Japandiscovered the fossils of Dipteridaceae floraClass Equisetineae (トクサ) Lobatannularia 30 m h. Neocalamites 2 m h (extinct in mid-Jurassic) Class Filicineae (シダ) Subclass Osmundidae Todites Subclass Leptosporangiatae (薄嚢シダ) Dipteridaceae Incertae sedis: Cladophlebis (form genus) Class Pteridospermopsida (シダ種子) Sagenopteris Class Cycadopsida (ソテツ) ☛ modern cycad Order Bennettitales: Otozamites, Pterophyllum Order Cycadales: Ctenis, Nilssonia Class Ginkgoopsida (イチョウ): Baiera, Ginkgoites, Czekanowskia Class Pinopsida: Podozamites, Incertae sedis (form genus) Class Incertae sedis Taeniopteris, taxonomical positions fiexed for a few specimens, not only from Japan
Class Eusporangiopsida: Marattiopsis (Marattiaceae) |
[Epoch: Jurassic 1.95-1.35 Bya]
|
Table. Geological distribution of Jurassic plants *: and allied species
Triassic
Jurassic
Cretaceous
Tertiary |
Seed plants in Mesozoic (中生代種子植物)Pollen (花粉)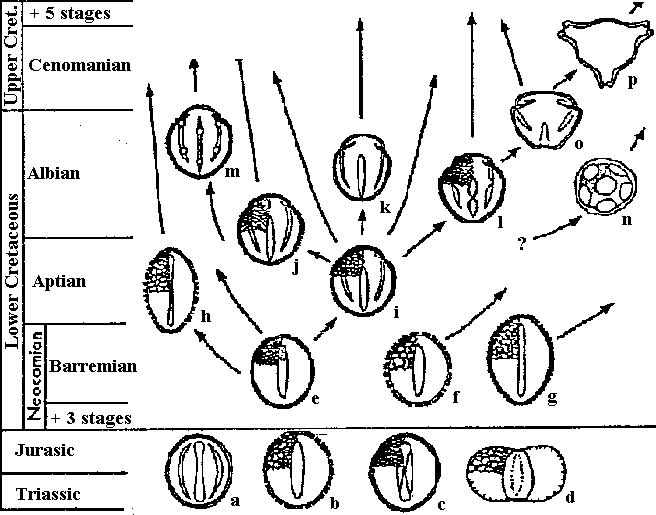 Fig. Time distribution and presumed relationships of principal early Cretaceous and Cenomanian angiosperm pollen types (e-p), and selected pre-Cretaceous pollen types (a-d). a: Eucommiidites, b: Triassic reticulate-columellar monosulcate of Cornet, c: cycad-type alveolar monosulcate, d: saccate alveolar pollen of Caytoniaceae and Corystospermaceae, e: Clavatipollenites, f: Retimonocolpites, g: Stellatopollis, h: Liliacidites, a possible monocot, I: reticulate tricoplate, j: striate tricoplate, k: smooth tricoplate, l: grain with tricolporate tendency, m: tricolpodiorate, n: polyporate, o: smooth, oblate-triangular tricolporate, p: early member of triporate Normapolles complex. LeafVein (葉脈): veinlet is present - developmental order of veins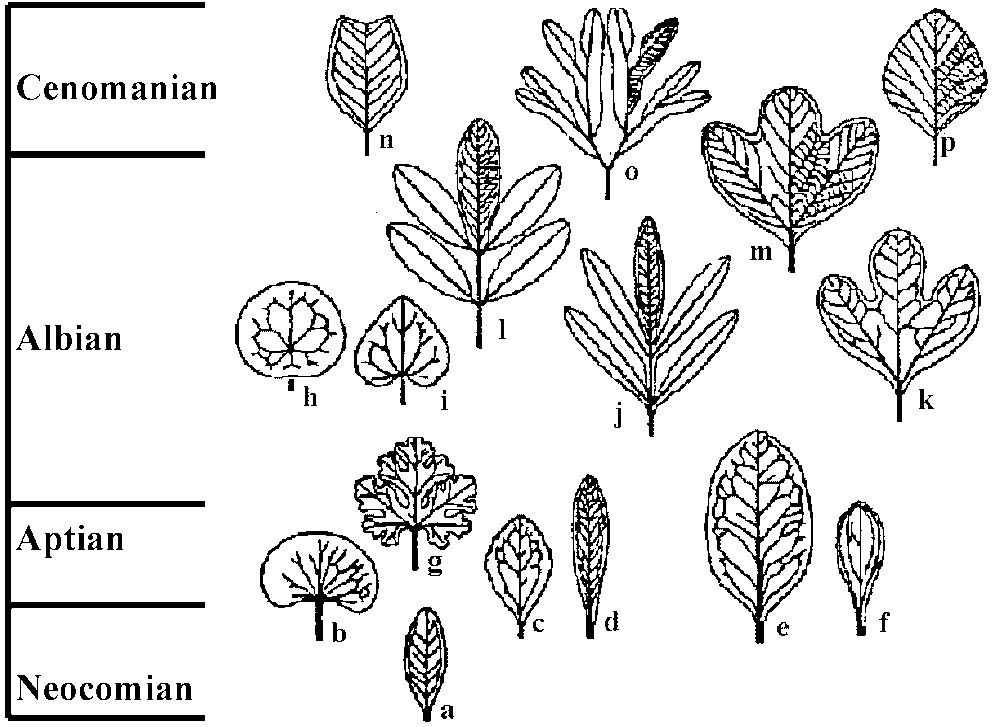 Fig. Principal early Cretaceous and Cenomanian angiosperm leaf types. a: small, pinnately veined leaf of Vakhrameev, b: reniform, c: serrate, d: oblanceolate, e: Ficophyllum, f: Acaciaephyllum, g: lobate reniform, h: peltate, actinodromous, i: ovate cordate, j: pinnatifid Sapinodopsis, k: early plantanoid, l: compound Sapindopsis, m: later plantanoid, with rigidly organized fine venation, n: Liriophyllum, o: dichotomously compound, p: secondarily simple platanoid derivative. |
[Epoch: Cretaceous 1.4-0.65 Bya]
|
Table. Pre-Aptian (≈ early Cretaceous) angiosperm fossil record → first angyosperms Age: Megafossil claims (wood, leaf, etc.) ↔ Pollen claims Barremian: Onoana ↔ Clavatipollenites Hauterivian: Valanginian: Carpolithus Barriasian: Tyrma fruits ↔ Tricolpollemites 'Tithonian': Problematospermum ↔ Pterocarya, Trifossapollenites Kimmeridgian: Palmoxylon, Ungeria ↔ Sporojuglandoidites |
Oxfordian: Sahnioxylon, Montsechia Callovian: Bathonian: Phyllites, Sogdiania ↔ Magnolia-type, Nelumbium-type Bajocian: Suevioxylon, Caytonia ↔ 'Clavatipollenites' Early Jurassic: Propalmophyllum, Gramiane, Sassendorfites, Fraxinopsis ↔ Euycommiidites, 'Clavatipollenites' Late Triassic: Furcula, Sanmiguelia ↔ EucommiiditesEarly Carboniferous: ↔ Tetraporina, Triporina |
Fish (魚類)[*: extinct] Coelacanth (シーラカンス)Coelacanthidae*: Axelia, Ticinepomis, Coelacanthus, WimaniaDiplocercidae*: Diplocercides Hadronectoridae*: Allenypterus, Hadronector, Polyosteorhynchus Mawsoniidae*: Alcoveria, Axelrodichthys, Chinlea, Diplurus, Mawsonia Miguashaiidae*: Miguashaia Latimeriidae: Holophagus*, Libys*, Macropoma*, Macropomoides*, Megacoelacanthus*, Latimeria (present), Undina* Laugiidae*: Coccoderma, Laugia Rhabdodermatidae*: Caridosuctor, Rhabdoderma, Whiteiidae, Whiteia Dinosaur (恐竜) Dinosauria Owen 1842: a type of reptile that became extinct about 65,000,000 years ago. There were many different types of dinosaur, some of which were extremely large. (general def.). 1: Brachiosaurus, body length = 25 m, height = 16 m. 2: Diplodocus, bl = 20-33 m, 3: Iguanodon, bl = 10 m, 4: Tyranosaurus, bl = 11-13 m, 5: Stegosaurus, 6: Triceratops, bl = 8-9 m, 7: Velociraptor, bl = 2 m, 8: Compsognathus, bl ≈ 1 m
Dinosauria Owen 1842: a type of reptile that became extinct about 65,000,000 years ago. There were many different types of dinosaur, some of which were extremely large. (general def.). 1: Brachiosaurus, body length = 25 m, height = 16 m. 2: Diplodocus, bl = 20-33 m, 3: Iguanodon, bl = 10 m, 4: Tyranosaurus, bl = 11-13 m, 5: Stegosaurus, 6: Triceratops, bl = 8-9 m, 7: Velociraptor, bl = 2 m, 8: Compsognathus, bl ≈ 1 mMammal
origin of living placental and marsupial mammals
15 cm l including tail, insectivorous (earthworm, etc.) Elephant (ゾウ)Elephantidae, elephants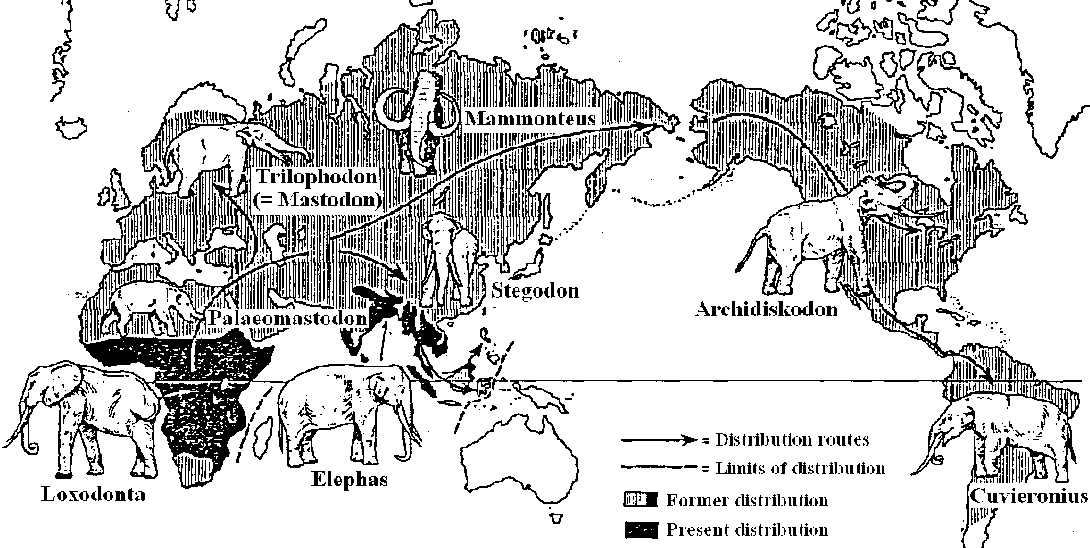 Fig. 83. Former and present distribution of Proboscideans: in the Late Tertiary natives of Africa, Eurasia, and North America; in the Pleistocene in South Africa as well. Disjunctive distribution of modern times (Africa: Loxodonta africana; South Asia: Elephas maximus). In the Late Tertiary and Quaternary spreading over the Arabian peninsula, Berign and Panama bridges. |
Primates (霊長類)1837 Lartet, Édouard: Pliopithecus antiquus in Europe and Asia
17-7 Mya, origin of gibbon
12.5-11.1 Mya in French Pyrenees 1862 Prof. Cocchi, Igino: fossils in a lignite mine at Montebamboli, Italy O. bambolii 1906 Osborn: identified an extinct primates, Apidium phiomense
theeth in early Oligocene (30-28 Mya) in the Fayoum deposits of Egypt 1911 Schlosser: identified Propliopithecus haeckeli
1963-64 many fossils by the intensive investigation of Yale Univ
1909 A gold prospector discovered the fossil
neither adapiform nor omomyid primates Pondaungia cotteri 1962 Simons: a primate, Oligopithecus savagei
- discovered in early Oligocene in Africa in Jebel Qatrani Formation, Egypt in 33.3-35.4 Mya Paleocene in Tertiary (第三紀) or earlier70 Mya: a primate-like animal, Purgatorius ceratops in Montana consisting of a single tooth found in late-Cretaceous rocks (controversial) 56 Mya: a mouse-sized Teilhardina brandti in Wyomingfingers curled around a branch [The origin of human deomonstrated by fossils ]Gigantopithecus von Koenigswald 19352-0.3 Mya (early–middle Pleistocene)1915 Pilgrim: described G. giganteus moved to Indopithecus giganteus 1935 von Koenigswald: described G. blacki, Black Eye
characterized by the huge teeth (morphologically human-like) 1945 Weidenreich: G. blacki grouped into Hominidae ⇒ Giant hypothesis (rejected now) 1956 Pei et al.: > 1000 fossils recovered from a cave in SE China
middle Pleistocene (0.75-0.5 Mya) overlapped with Homo erectus grouped into Dryopithecus or Pongidae 1969 Simons & Chopra: G. bilaspurensis in Bilaspur, north India
in the Pliocene of Tertiary (9-5 Mya) |
☛ Epoch: Cainozoic
Tertiary (第三紀), 64.3-2.6 MyaPalaeogene, Paleo-tertiary (古第三紀)Insects diversedPrimitive mammals developed
Nummulites, nummulite (貨幣石) Neogene, Neo-tertiary (新第三紀) 25-2 MyaModern mammal development [warming → cooling] |
Animals:
Lepidocyclina - foraminiferan Miocene (中新世) 23-5 MyaMammals developedPliocene (鮮新世) 5-2.58(1.6) MyaQuaternary (第四紀) > 2.6 Mya-presentPleistocene (更新世) 2600 Tya (2.6 Mya)-11.5 TyaHolocene (完新世) 11.5 Tya-Present |
Migration region (habitat or biotype) and spread (移住と拡散)Regional patterns are decided by the indicator species
paleobiogeographical province acretion techtonics, land bridge (陸橋) + comparisons with the present biogeographical regions |
Relationships to Geology (地質学との関連)Biosparite: sparMicrite: lime mud Adaptation spar: trace fossil Paleoequator (古赤道)The position of the equator in the geologic past as defined for a specific geologic period and based on geologic evidence. When talking about plant distribution (by Maekawa), the specific period is between Cretaceous and Tertiary.Sedimentary ancient DNA (sedaDNA, 堆積物古代DNA) |
|
c. 1836 description of pollens in peat and coal by German scientists pollens discovered from brown coal Göppert, Johann Heinrich 1800-1884, (paleo-)botanist in Germany
coal - consisting of plant cells, confirmed by microscopic observations 1905: developed pollen profile by peat profiles in Sweden
established pollen analysis 1909 pollen diagrams of peat profile von Post, Ernst Jakob Lennart 1884-1951, Swedish geologist
1906 Linnaeus Prize, by the age estimation method of peat |
1916 "Forest tree pollen in south Swedish peat bog deposits", presented in 16th Convention of Scandinavian Naturalists in Kristiania (Oslo)
1909 "Postglaciala tidsbestämningar"
Pliocene: forests by temperate broad-leaved and needle-leaved trees
Pliocene - 2.6 Mya - Pleistocene (glacier period) 0.8 Mya: cool-temperate forests / subalpine needle-leaved forests |
= great extinction, mass extinction or biotic crisis (Raup & Sepkoski 1982)Big Five1) Ordovician-Silurian extinction event= End Ordovician (O-S)70% of species, 57% of genera and 27% of families were extinct 2) Late Devonian extinction70% of species, 50% of genera and 19% of families were extinct3) Permian-Triassic extinction event= End Permian, P/T extinction90-96% of species, 83% of genera and 57% of families were extinct |
4) Triassic-Jurassic extinction event= End Triassic48% of genera, 23% of families were extinct (55% of genera and 20% of families in marine) 5) Cretaceous-Paleogene extinction event= End Cretaceous, K-T extinction, or K-Pg extinction75% of species, 50% of genera and 17% of families were extinct ☛ endangered species / extinct species 6) And nowThe present extinction rate is 1000 times faster than the past rate, estimated by fossil recordThe future extinction rate is 10 times faster than the present rate |