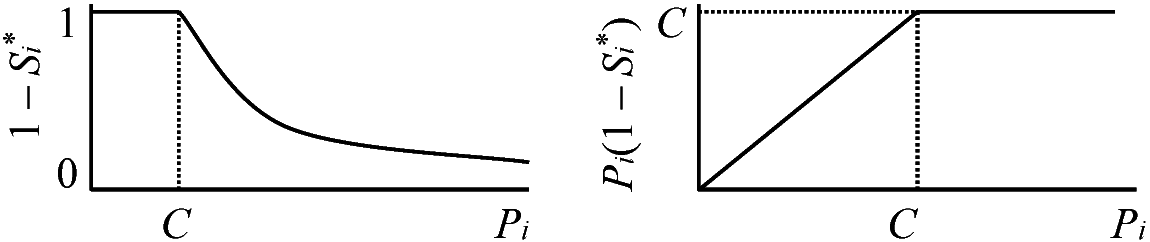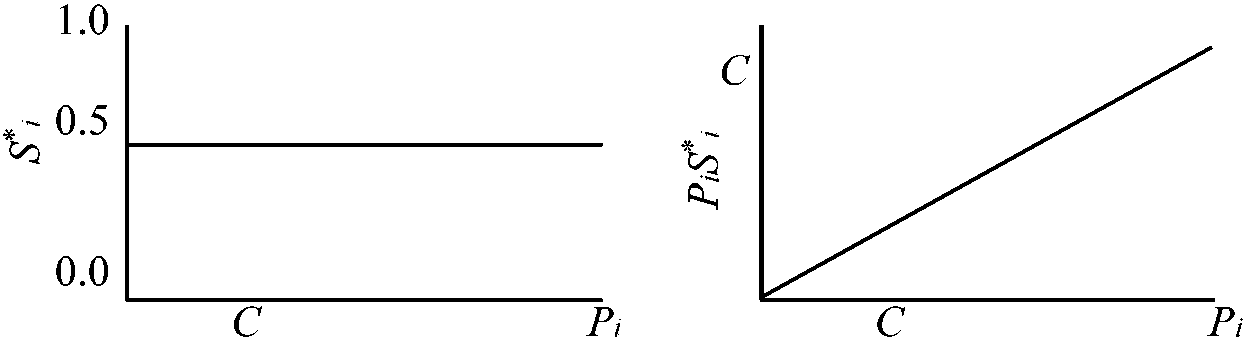(2026年6月20日更新) [ 日本語 | English ]
HOME > 講義・実習・演習一覧 / 研究概要 > 小辞典 > 生活史
|
≈ 生活環 life cycle (生態学では) 個体の出生から死亡に至るまでの過程 → 生活様式に関する諸問題(環境・形態・生理等)を軸に生物の一生を表現 植物は、種子から発芽定着することにより生活史が始まると考えがちだが、生活史は生活環と言われるように1つのサイクル(life cycle)を有する。 Ex. 植物が最も移動できるときは花粉と種子の段階であり、動物でいえば種子は大人である。生理生態の解明を目的とするならば、次世代への適応戦略を頭に置くことが必要 世代 generation世代単位: if (無性繁殖除けば)個体の有する遺伝子に変化が起こった時 → 種子(またはそれ以前の胚の発生段階)が新世代の始まり ☛ 蘚類 | 羊歯類☛ 生活型 鶏が先か卵が先か: 生活環はどこから初めてもよい。ただし、個体の始まりを起点とするなら卵から |
生活史段階実生 seedling発芽-成熟までの期間 → 植物生活系(一年生・多年生・木本等)により具体的期間は異なる繁殖までの成長様式が重要であり、関与する環境要因・競争関係の調査が繁殖成功測定にに必要 1932 Boysen-Jensen: 実生のジレンマ成長最適環境と発芽最適環境は異なることがある → 自分の生存により有利な生息地を選ぶならどちらかを犠牲にせねばならない(Aguiar et al. 1992) 1970 Janzen: 母樹からの距離と実生数(再加入数)の理論的モデル木の近くでは全種子が種子食動物に食べられるが、木から離れるに従って摂食者減り、種子生存確率が増す。母樹からの距離に従って散布される種子数は減少するにもかかわらず、再加入率は木からある距離離れた地点で最大になる |
グライムの三角形 Grime's triangle (☛ 例)耐性(stress tolerance) vs 撹乱(disturbance) vs 競争(competitive ability)一年生植物 annuals種子から種子までの生活を1年以内で終える草本植物 - したがって越年生は1年生に含まれる
夏型 1991 Barbieer et al.: 野生イネ2種16系統 乾燥地域において多年生 → 1年生進化 2005 Evans: Oenotheraの21種を種間比較降水量の少ない生息地に1年生草本が多い 開花のタイミングQ. 繁殖活動と栄養器官成長との間で、光合成獲得物質をどのようなスケジュールで分配するのが最適か |
A._
dx/dt = (1 – u(t))g(x)
dy/dt = u(t)g(x)
x: 栄養器官のサイズ 多年生植物 perennials2年以上にわたって生育する植物 = 一年生・二年生植物以外の全て宿根草 (≈ perennial)ストレスの高い時期(冬期や乾燥期)に、地上部を枯らし地下部で生存する戦略をとる多年草宿根: 多年生植物の地下部 |
|
季節変化に伴う動植物の行動・状態・形質等の変化
Ex. 発芽, 開葉, 開花, 紅葉, 落葉 光: 太陽光変化に影響される生理的・生態的過程 (Chazdon 1988)1) 時間スケール
Ex. 光周性 + 生物時計(体内時計 Ex. 日周期 diurnal, or サーカディアンリズム)
|
春植物 (spring ephemeral)= 早春季植物春先に植物群集上層部の葉が展開し光量が急に低下する前に葉を広げ、開花結実し、光量が低下時には地上部を枯らし休眠状態に入る (例, examples) 熱呼吸= 開花時寒冷対策
春、花芽成長時に数cmの雪を被るが、発熱で融雪し氷雪にトンネルを掘り、花軸が伸び、その上につく花芽が雪上に出て開花する 展葉落葉性植物の葉の開葉・落葉戦略は2タイプに分けられる(菊沢 1986)
一斉展葉タイプ: 極相林など安定な場所
個体小 → 遅くまで葉を展開 ⇒ 大きくなるにつれ次第に展葉早めるよう変化(予測) 花の寿命 longevity of individual flowers他殖率(c)推定式(Primack 1995): 仮定 = 直接開花数が花寿命に影響するc = 1/(fp)
c: 他殖率 outcrossing rate
c = 1/(3 × 10) = 0.033 ≈ 3% |
[ 繁殖/生殖 ]
性/雌雄 sex (female and male)雌(♀): 大型で養分を多量に含む配偶子作る個体 – 初期発生を賄う雄(♂): 小型で運動力に優れた配偶子作る個体 – 養分殆どなし 性型 sexual formCase. 植物雌雄同株(単性雌雄同株・雌雄異花同株・雌雄混同株・雑居花・雑性花) hermaphrodite and monoecy (synoecious): ♂♀配偶子が1個体に作られる= 両性花と雌花又は雄花 male flower (staminate flower)が同一株につく
雄性先熟 protandry
雌雄同熟花 adichogamous flower 装飾花(不稔花) ornamental flower: 特に大型で美しい Ex. ガクアジサイ、ヤブデマリの装飾花 雌雄異花同株 monoecism, or monoecy: 単性花を形成 (♂/♀)単性花 unisexual flower: ♂f = 雄蕊、♀f = 雌蕊 → 一方のみ有する Ex. カボチャ、キュウリ、スイカ 雌雄同花序 androgynous: 花序に雄花・雌花が混在するもの 三性同株(雄性雌性両全性同株) androgynomonoecious, trimonoecisous: (♂/♀/♂♀)雄性同株(雄性両全性同株・雄花両性花同株) andromonoecious: (♂/♂♀) 雌性同株(雌性両全性同株・雌花両性花同株) gynomonoecious: (♀/♂♀) 雌雄異株 dioecism, dioecy: 雌性配偶子個体(雌株) + 雄性配偶子個体(雄株)(♂)/(♀) Ex. ホウレンソウ、Morus, Ginkgo、ソテツ、スギゴケ、ゼニゴケ
三性異株(雄性雌性両全性異株) androgynodioecious, triecious: (♂)/(♀)/(♂♀)
雌株は花粉を作らない分、より多くの種子を生産できる (不完全雌雄異株 subdioecy, 雌雄混株のsubdioecious) 多系(雌雄混株・雑居性雌雄異株) polygamous, polygamny: 離性内の不完全雌雄異株のうち雄性異株・雌性異株を除いたもの
雑居同株 polygamomonoecious: 多性 polyecy (adj. polyecious)が一部見られる不完全な雌雄同株 植物での進化仮説自家交配回避(近交弱勢回避)花粉補償 捕食者回避 ディスプレー (雄花) Def. 近交弱勢 inbreeding depression: 近親交配 → 子孫に不利な形質 Ex. 健康低下、遺伝的異常・疾患増加、繁殖の低下、寿命短縮 Case. 動物雌雄同体 hermaphrodite: 1個体に卵巣と精巣を持ち、卵と精子を作る動物(普通卵成熟期と精子成熟期が異なり自家生殖起こらない) Ex. ミミズ、マイマイ、サナダムシ、ジストマ、ホヤ常時雌雄同体 simultaneous hermaphrodite
同一個体が同時に卵巣と精巣(雌雄生殖機能)もつ生物
一生中で性が変わる生物 Ex. 雌 → 雄 雄 → 雌
Ex. カキ: 繁殖期後に中性状 ⇒ 栄養状態良 → ♀(卵作れる) 不良 → ♂ 性決定 sex determination☛ 細胞遺伝学 性染色体 sex chromosome環境 environment雌ホルモン, 酵素, 年齢, 水温等
性転換 sex transformation社会構造・婚姻形態に関係Ex. スズメダイ: 雌雄異体 - 性転換なし Case. 雌性先熟: ♀ 体サイズ ∝ 卵生産数 ♂ あるサイズより大きいと♀独占 ♀ if 体サイズ ∝ 齢 ⇒ 成長と共に繁殖成功度増加 ♂ 大きな♂は♀独占 - 小さい♂は繁殖できない 小さい♀繁殖成功度 ≈ 0 ⇒ あるサイズから♀独占 ∴ あるサイズで急激に繁殖成功度増加 性転換が繁殖成功に有利 ⇒ 一夫多妻進化(雌性先熟有利) Case. 雄性先熟 Ex. クマノミ: イソギンチャク = 生息地(他への移動困難)
1イソギンチャクに1-数匹のクマノミ
♀ 体サイズ ∝ 繁殖価 vs ♂ 体サイズに関わらず繁殖価一定
第一位(♂)が死ぬと第二位が卵細胞成熟させ♀となる 密集状態 → 一夫二妻・二夫一妻生じるが一時的なもので不安定 産卵: イソギンチャクの近くの岩盤 - ♂が卵の世話 ♂を除去すると♀が卵の世話をする
孵化: 産卵10日後 - 稚魚: ≈ 2週間浮遊生活 → イソギンチャクに入る
性転換 = 大きなコスト必要 - クマノミでは性転換に1-2ヶ月必要 ⇒ 性的単形と多形 sex monomorphism and dimorphism1871 Darwin: 性区別が一見でわかる生物とそうでない生物がある
性的2形dimorphismのある動物は大体一夫多妻 polygamy 大型哺乳類一腹産子数 litter size = F1(仔数)/一腹: 哺乳類は他と比べて小妊娠期間長い long gestation period → 仔数/♀= 次世代数は母親かF1数により決定される 一夫一妻ならF1遺伝子は限定的Ex. ヒト = 一夫一妻 → 有害遺伝子増加(Ex. フェニケルトン症) 一夫多妻: 限られた♂だけが遺伝子残す= 環境により適応した遺伝子を有する♂だけの遺伝子が残る 単形の場合Ex. 小型哺乳類: 集団サイズpopulation size大 → F1で淘汰されてもsizeが大きいので問題無鳥: 一夫一妻多 雄同志の争いは飛翔不可能にしがち → 自然にmonogamy発生と考えるのが妥当 ← polygynousだと雄同志の闘争激化 性淘汰 (sexual selection)1) 個体の最適性比 optimal sex ratioa. intrasexual 同性個体間(の競争)要素b. epigamic 異性間の要素 - より良い配偶者を選ぶ競争 1956 Maynard-Smith
ショウジョウバエではhomogamic, heterogamicの交配結果としてheterogamicの方がhomogamicの4倍の孵化率を示し孵化率が全く異なった 2) 配偶システム(婚姻形態) mating system繁殖構造 breeding structure同型交配(配偶) homogamy: 同じ形質持つ個体が交配 同類交配 assortative mating: 類似表現型個体同士がより頻繁に交配 異型交配(配偶) heterogamy: 異なる形質持つ個体が交配交配様式 mating system a. 単婚(一夫一妻) monogamy: 資源防衛、♀防御、♂優劣関係 Ex. 多くの小鳥、ツル、テナガザル 魚類: 一回の子育てが終了するまで両親とも子の側に留まる (淡水魚に多)同じ一匹の相手のみと繰り返し繁殖する (海水魚に多) b. 複婚 polygamy一夫多妻 polygyny Ex. コウライキジ、マントヒヒ
経時一夫多妻 polybrachygyny Ex. アシカ、オットセイ、ライチョウ 多夫一妻 polyandry Ex. オオヨシキリ、レンカク類、クイナ類の一部 c. 乱婚 promisquity Ex. ニホンザル(稀, 普通ランダムではない)下等脊椎動物で複数卵に放精されることもあるが化学的に拒否される |
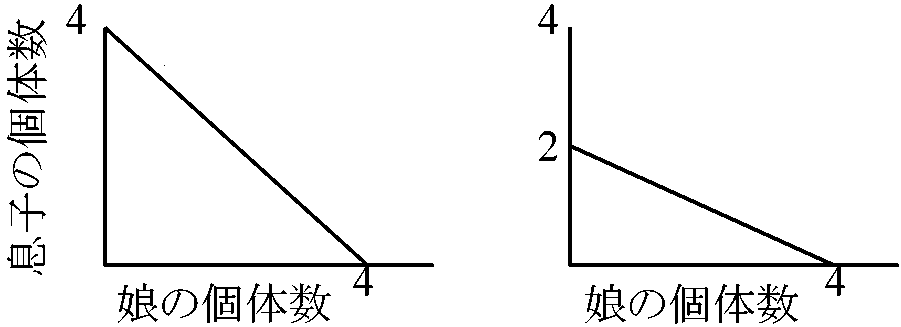
性比 sex ratio性比進化: 子を作るのに使える資源量(子への総投資量)の娘-息子配分比仮定: 性比に遺伝的基盤存在 = 自然選択の対象 → 個体の適応度を最大化する性比進化
(a) 性的2型がない (b) 性的2型がある 性比モデル総投資量の個体変異を考慮した場合における個体の最適性比の予測1)母親が子供の性比調節能力を持つことを仮定し、 2)繁殖集団構造を4つに分類し、 3)子への総投資量の個体変異を組込んだモデルを作る 繁殖集団patchを創設する母親数をnとすると母親iの包括適応度は Fi ∝ rdPi(1 - Si)Rf + rsPiSiRm (i = 1, 2, …, n) … (1) (Fischer式)
Pi: 母親iの子への投資量, Si: 母親iの性比(投資量の性比) Rm(雄の単位投資量あたりの繁殖成功度)について a) 雄は自分が生まれたパッチに留まり、その中で無作為交配 Rm = [Σj=1nPj(1 – Sj)]/[Σj=1nPjSj] × Rf b) 雄は交配前に分散しランダムにパッチに入り、その中で無作為交配
Rm = constant a) 雌は交配後もパッチ内に留まり、資源をめぐって雌同士で競争
Rf = K/[Σj=1nPj(1 – Sj)] b) 雌は交配後分散し、ランダムに新しいパッチを作る Rf = constant 表. 以上を組合わせると、繁殖集団構造を4分類できる
♀/♂
A
| B
LMC (local mate competition): 雌をめぐる雄同士の局所的な競争 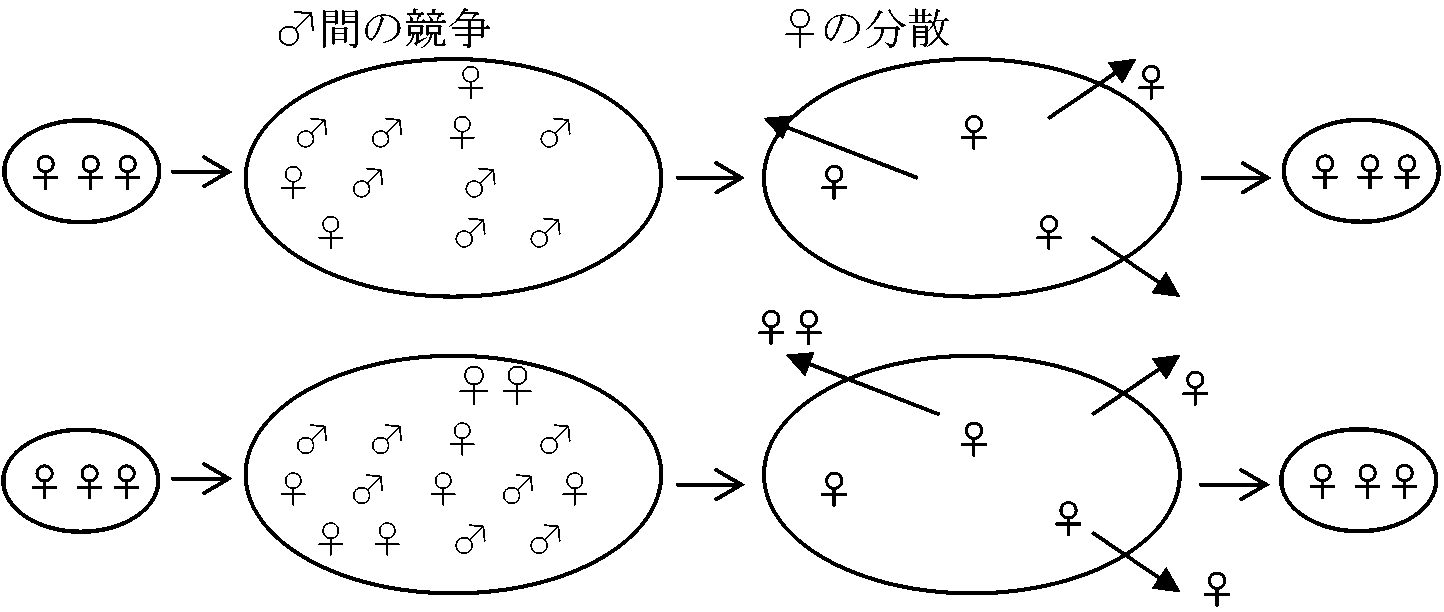
Fi ∝ Pi(1 – Si)Rf + PiSi{ΣPj(1 – Sj)/ΣjnPjSj}Rj … (2)
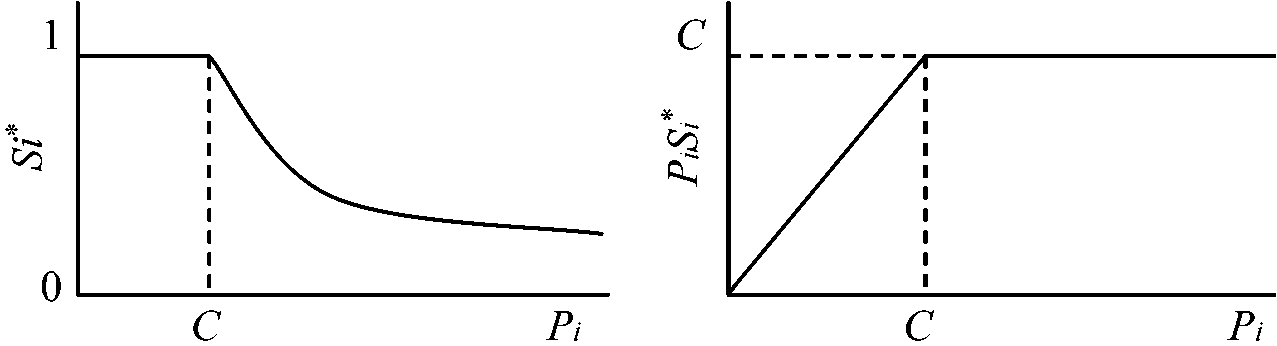
Si* = 1 for Pi ≤ C (i = 1, 2, …, m) … (2a)
C = {S-(1 – 2S-)ΣPi}/Pi = [(n – m – 1) + √{(n – m – 1)2 + 8(n – m)ΣmPi/ΣnPi}]/{4/(n – m)}
(2b) → PiSi* = C 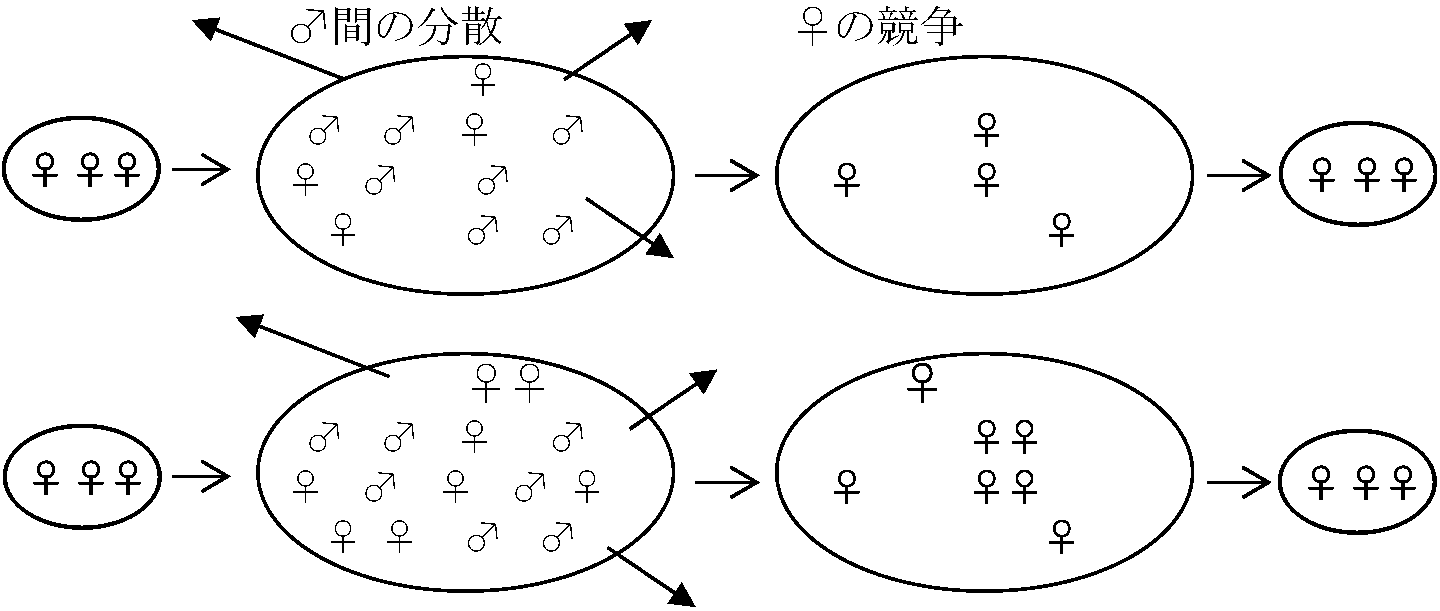
Fi ∝ Pi(1 – Si){K/Σj=1nPj(1 – Sj)} + PiSiRm … (3) 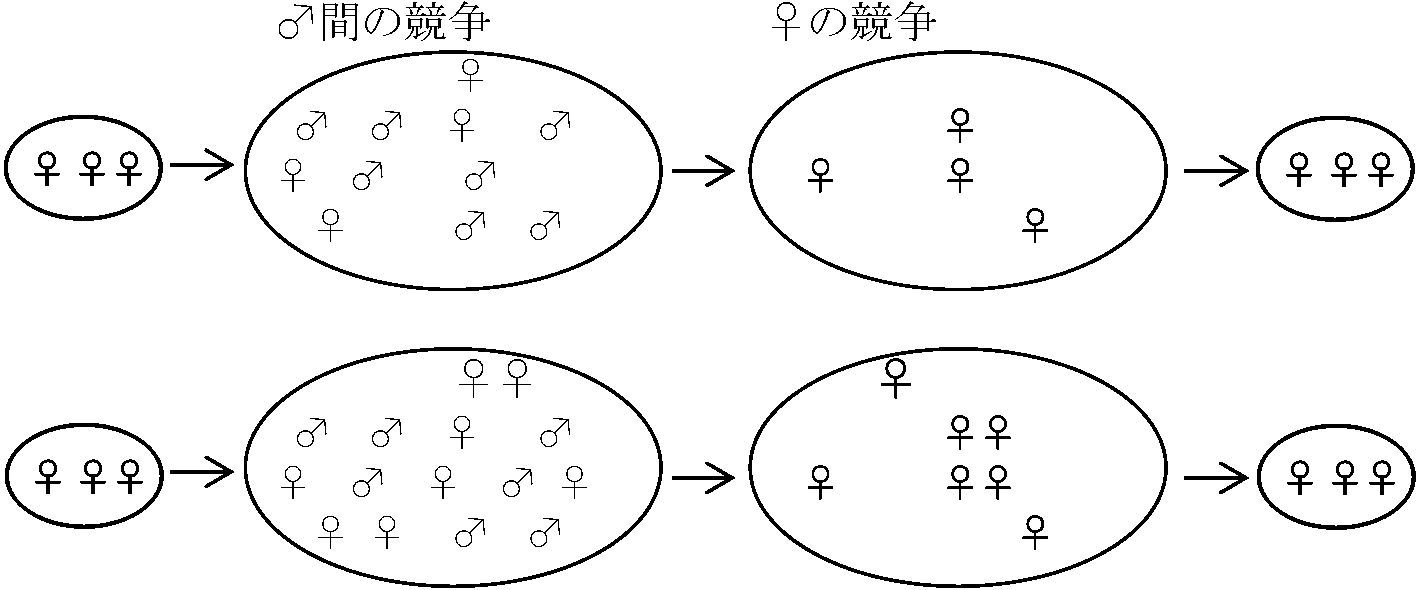
Fi ∝ Pi(1 – Si)·K/{Σj=1nPj(1 – Sj)} + PiSi·{Σj=1nPj(1 – Sj)/Σj=1nPjSj)}·{K/Σj=1nPj(1 – Sj)}
Si* = 1/2 「総投資量に関わらず、どの母親も1対1の性比で子供を作る」 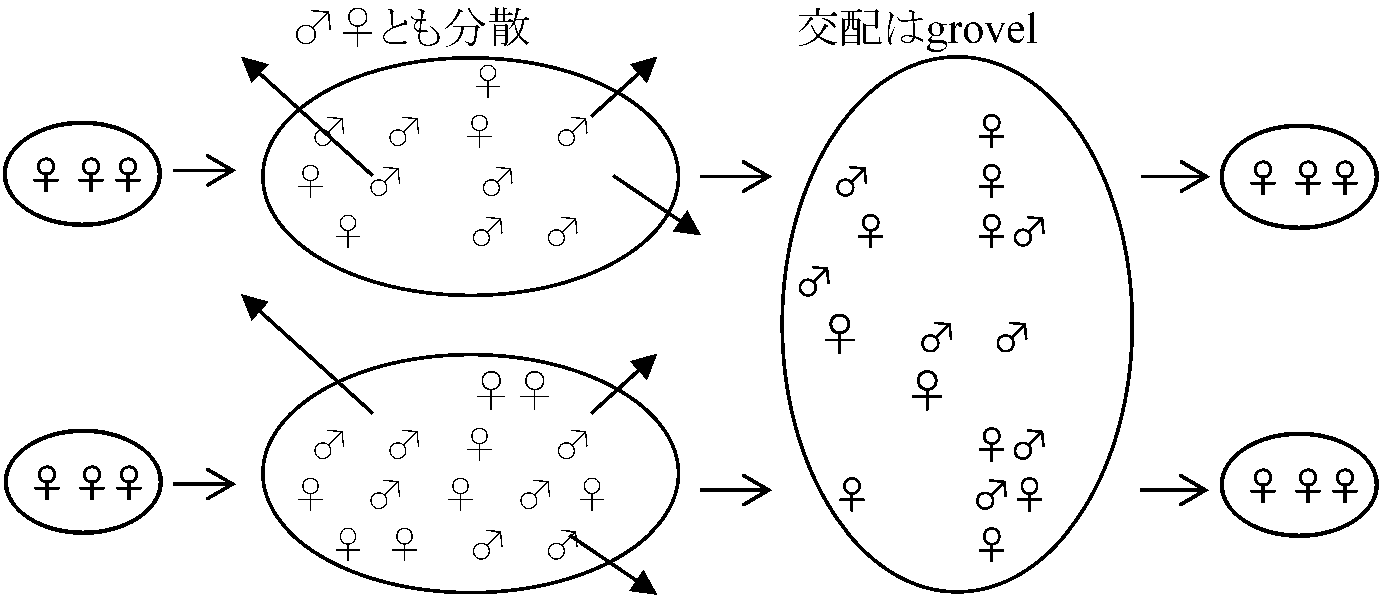 |