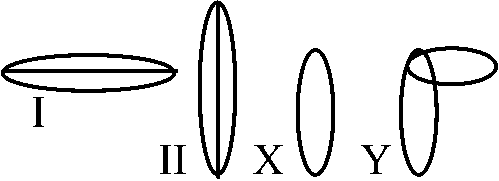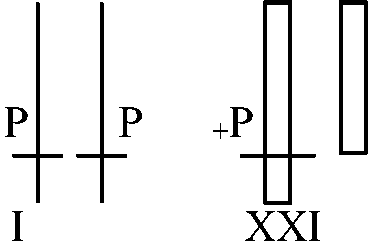(2026年7月27日更新) [ 日本語 | English ]
HOME > 講義・実習・演習一覧 / 研究概要 > 小辞典 > 生物学 > 遺伝学
1900 de Vries, Correns, Tschermak: メンデルの法則再発見 ⇒ ダーウィン派 vs メンデル派 - (感情的)対立 ダーウィン派: 微小連続変異に淘汰が働き進化 1902 Weldon WFR: 安定化(求心性)淘汰発見 - 呼称は(Fisher 1930)
巻貝死亡率 - 巻き数が平均的なもので低 = (後の)安定化淘汰 Bateson, William (1861-1926): 遺伝学(genetics)用語提唱 (1906)
1894 Materials for the study of variation: 進化に不連続変異重要 |
1903 Johansen W (1857-1927): 純系説 - 純系に淘汰働かない 1908 Hardy HO, Weinberg WW: ハーディ・ワインベルグの法則 1926 Morgan TH: 遺伝子説 - 矛盾なく対立解決することを試みる 1927 Muller HJ: 突然変異人為的に誘導 - 突然変異説支持 1930 Fisher RA: 自然淘汰の遺伝学的理論
自然淘汰 ≠ 進化 ⇒ 自然淘汰の問題を進化から切り離し独自分野とし研究すべき |
|
( メンデル 1865) メンデルの法則 (Mendelian rules or laws)メンデルの実験 Mendel's experiment材料: エンドウ – 誰でも明らかに区別できる7形質選び交配実験
対立形質__ 対立遺伝子 allele 体細胞上に遺伝子は2個づつあり、生殖細胞には1つある 優性の法則 dominance law形質異なる親が支配できるF1には一方の形質のみ表れる(適用できない例多)F1 = Aa ⇒ F2の表れ方 ♂♀ A a
Parents 〇AA [A(生殖細胞)] × ☆aa [a] → 3 (AA + Aa + Aa) : 1 (aa) 雄蕊 stamen、雌蕊両方に上の表の確率で生殖細胞が作られる→ 子は3:1で表れる(分離の法則) → 実験結果を良く説明 分離の法則 segregation lawF1生殖細胞形成の際、Pから来た対立形質を示す遺伝子が、それぞれ分離し別々の細胞に均等配分 → 最初の父方と母方の形質が分離し表れる ⇒体細胞には遺伝子が2個あり、生殖細胞形成時に1個づつ分離されるため 表現型 phenotype: 実際に表れる形質。Ex. 種子形 → 丸/皺 → フェノミクス(網羅的表現型解析) phenomics 遺伝子型 genotype: アルファベットの並び方 Ex. AA = 優性ホモ homo、aa = 劣性ホモ、Aa = ヘテロ hetero 独立の法則 independent-assortment law各形質は他の対立形質に左右されず独立に遺伝Ex. 丸黄 角黄 丸緑 角緑 計
AB Ab aB ab
丸:角 = (315 + 108):(101 + 32) = 423:133 ≈ 3:1 = (9 + 3):(3 + 1) Ex. エンドウ種子色: Y gene = 黄/緑 + R gene = 丸/皺 Y/R = 非対立遺伝子 ⇒ 「独立の法則」典型 単性雑種の応用1) 不完全優性(中間雑種): 不完全優性 = Aとaの差がない
Ex. Ipomoea purpurea花色
Ex. ハツカネズミ体色
A: 黄色遺伝子。ホモ(AA) = 劣性致死遺伝子 劣性致死遺伝子: ヘテロ接合状態で個体に保持される 1956 Morton JF, Crow JF & Muller HJ: ヒト1.5-2.5個の致死相当量lethal equivalent劣性遺伝子(個体では倍) 1958 Crow JF: 遺伝的荷重 genetic load, L ≡ 遺伝子型レベルで働く淘汰量集団中に含まれる最適な遺伝子型(wop)と比べ平均淘汰値(集団の平均適応度, wav)が低下する率 L = (wop - wav)/wop Ex. 正常野生型遺伝子(+) vs 劣性致死遺伝子(l), 突然変異率 := v →q^ = v/h (q^ ≡ 致死遺伝子lの平衡状態での頻度 h ≡ 有害度)
+ + 最適遺伝子型: 最適適応度, wop := 1 → 頻度 (1 - q^)2 集団での平均淘汰値: wav = 1·(1 - q^)2 + (1 - h)2q^(1 - q^) ≈ 1 - 2hq^
∴ wav = 1 - 2v (∵ q^ = v/h) ⇒ 有害度(h)に依存しない (前提: h ≠ 0. h ≫ v) [ 血液の機能 ] (古畑 1962) 血液型 blood type3) ABO式血液型遺伝 ABO blood type: 1対立遺伝子に3種類の遺伝子
(A, B, O)関与 = 複対立遺伝子
異種血球凝着反応: 異種動物血液を混ぜると凝着する 凝着反応は[A - α] or [B - β]共存時 ⇒ 3型(A, B, C)発見 (C: 後の0) |
血液型 A B AB O → 血液型には凝着原の名前がつけられた 1902 von DeCastello and Sturli: 3型に属さない型確認 = 現在のAB型判定法: 標準血清 凝集反応 A型血清(βを持つ): 抗B血清 = 黄色 – + + – B型血清(αを持つ): 抗A血清 = 青色 + – + – 血液型 A B AB O Q. 抗A型血清反応+ = 12人、抗B型反応+ = 20、両方+か– = 16 → 人数 A. 各血液型数: A型+B = 12, B+AB = 20, AB+O = 16, A+B+AB+O = 40 A = 16(人), B = 8, AB = 4, O = 12
↗ AB ↖ ↔AB
◎ 輸血血液の凝着素(血清中)と受血者の凝着原(血球上)で凝着する時 Ex. カエデ: O型 = 紅葉(アントシアニン) vs AB型 = 黄葉(カロテノイド) 色に合わせて血液型も変化: AB型 → O型 Ex. B型: アセビ、イヌツゲ、セロリ、ツルマサキRh式血液型 ☛ 輸血学 1937 Landsteiner: Macaca mulatta赤血球をウサギに接種し抗体を作らせる 1940 Landsteiner & Weiner: その抗体(抗Rh式血清)を利用し調査 抗Rh血液 → M. mulatta血液: 凝着(+) → ヒト血液(+ or -) 凝着(+) = Rh+型 → Rh因子Rh factor (M. mulattaと共通起源)持つ ↔ (–) = Rh–型 → 持たない その他の血液型: 犯罪証明、親子鑑定等に利用
MN式、Q式、E式等 両性雑種dihybridの応用両性遺伝: 遺伝子の相互作用1) 補足遺伝互いに補足的に働く遺伝子
P AABB (violet) × aabb (white) Ex. スイートピー花色
A: 紫色色素を作る遺伝子
a: 紫色色素を作れない
RRpp (rose) × rrPP (pea) Ex. ニワトリのトサカの遺伝
R: バラ冠を作る遺伝子 ⇔ r: 単冠となる 2) 抑制遺伝 inhibitor
P AABB (white) × aabb (white) Ex. カイコの繭色
A: 色が作られるのを抑える遺伝子(抑制遺伝子) 3) 同義遺伝 multiple gene system同形質を支配する2対以上の対立遺伝子があり、そのいずれか一つの遺伝子が優性であれば、優性の形質を示す遺伝現象がある。このような遺伝子
P ウチワ型 T1t1T2t2 Ex. ナズナの果実形
T1, T2: 団扇型遺伝子 ⇔ t1, t2: 槍型遺伝子 4) 条件遺伝
P CCGG (grey) × ccgg (white) Ex. カイウサギの毛色
C: 着色遺伝子(= 条件遺伝子)
c: 白色遺伝子 |
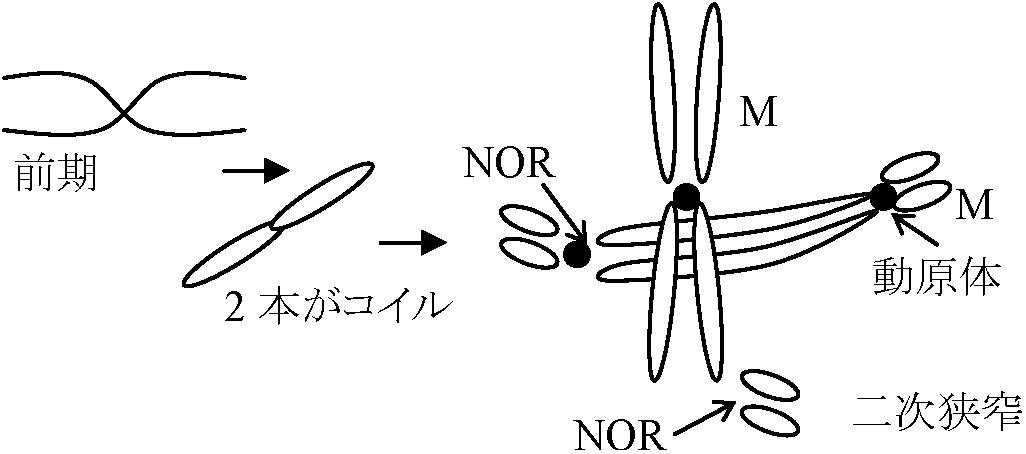
Def. 体細胞分裂 mitosis: 体形成細胞数増加し染色体数変化ない (2n → 2n)静止期(G1)体細胞分裂をしていない。物質交代が盛ん↓ 終期頃: 物質交代盛ん。DNA複製起こる(核が普通の細胞の倍になる) 分裂期(M) = 前期 + 中期 + 後期 + 終期前期: 核膜、仁消える。染色体表れる。紡錘体spindle作られる
植物: 極帽(動物の中心体に相当)表れ、紡錘糸張る
後期: 染色体が完全に縦裂
中期: 染色体が赤道面に並ぶして両極へ移動
染色分体の動原体 (centromere
紡錘体消える。染色体消える → 核膜、仁表れる 自己増殖系: ミトコンドリア、葉緑体などは自分で数を増やす Def. 減数分裂 meiosis: 生殖細胞形成時に染色体数は半分になる (2n → n)
半減 → 遺伝子組替の多様化 crossing-over (組替) 対合 pairing: 相同染色体の交差(乗換) crossing-overによる
分裂環 division cycle ⇔ 生活環 cell cycle
単細胞の場合には一致、多細胞の時には異なる
非分化: 増殖能力のある細胞 ⇔ 分化: 非増殖能力のある細胞
Hela cells, L-strain (mouse) 1907 Ehrich: マウス腫瘍細胞をマウス腹水中で増殖→ 何か本質的essentialなものを与えれば細胞は延命する 1940 カレル: ニワトリ胚で同様な結果1961 ヘイフリック
4ヶ月の胚の肺から組織を取り出し細胞を観察 → 50回分裂後死亡 カルス callus分化キャンセル → 再び増殖可能 → ニンジン組織培養(脱分化)オーキシンにより制御 = オーキシンを制限すると1個体になる Radioautography: Strip method, Dipping method1955, 56 Mazia & Plaut: 14C-thymidineを始めて使用 (現在は3Hが一般的)
DNA合成素材 (N-bases, i.e., A, G, C, T, phosphate, deoxyribose)
DNA合成が間期interphaseに起こることを示す 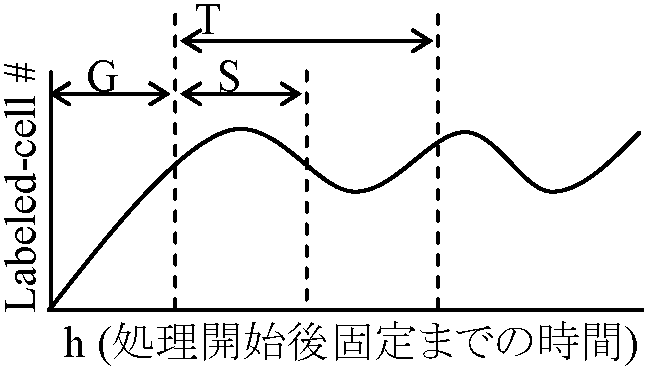 1964 Evans & Scott: Vicia fava
1964 Evans & Scott: Vicia fava
細胞周期決定
染色体レベルの複製様式
DNA合成後のmeiosisで3H-TdR → 3H-TMPとなるかどうか(thymidine kinase活性)を調べる heterochromatinは最初にできる例を発見 1966 Brown: 染色体がcondensedするとinactiveになる → single activeで可
条件次第で2本のどちらがcondensedしinactiveになる 1971 Summer & Carnoy: SSC (acetic acid/SSC/Gimsa) → G-band 核分裂 nuclear division or karyokinesis と細胞質分裂 cytoplasmic division or cytokinesis一連の分裂過程だが全く別の現象とも捉えられる - 多核現象・重複受精内胚乳 endosperm で分裂はsynchronizedしている 前減数と後減数一般に前減数であるが、Paspalumなどで後減数が見られる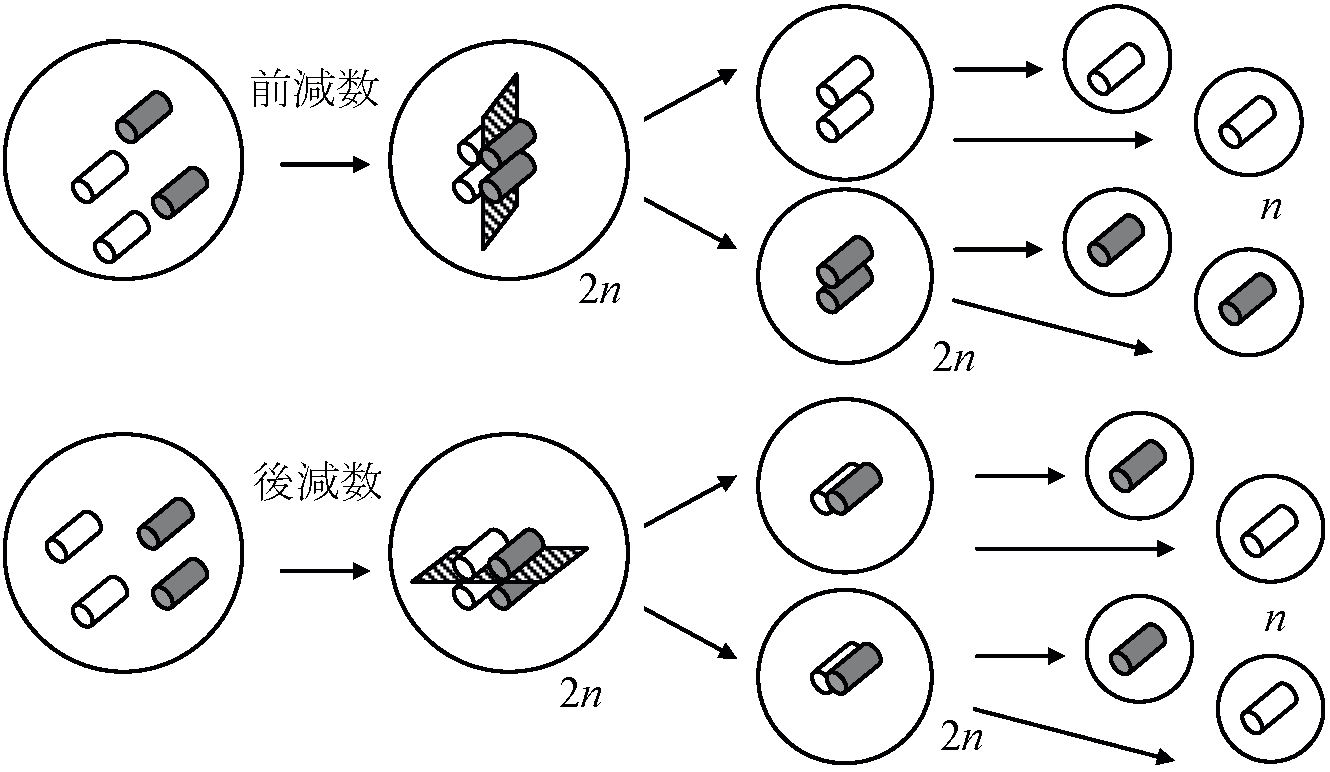
|
相 phase
metaphaseでchromosomeは1本に見え、その前後では対をなして見える。これは2本のchromosomeがうまくcoilしているためと考えられる。核小体は細胞質中にあったものが再発現 核小体が集中しよく染色しない
--------------------------------- 5S r-RNA
45Sが切れ18S, 28Sになる途中のものは全てimmature RNA 一価染色体の行動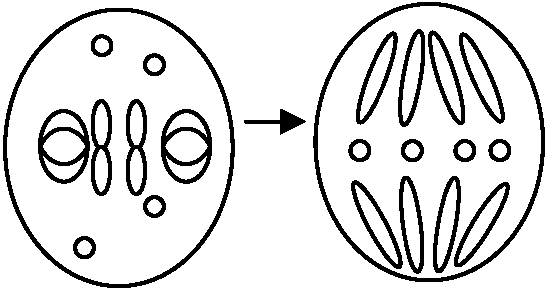 3通り確認された
3通り確認された赤道板外に配列(第I分裂中期) → 第II(I)分裂後期 任意分配あるいは縦裂分裂
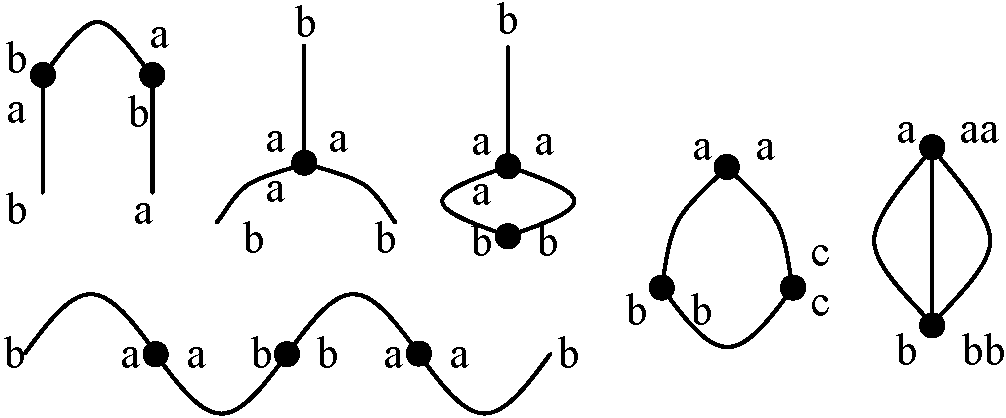
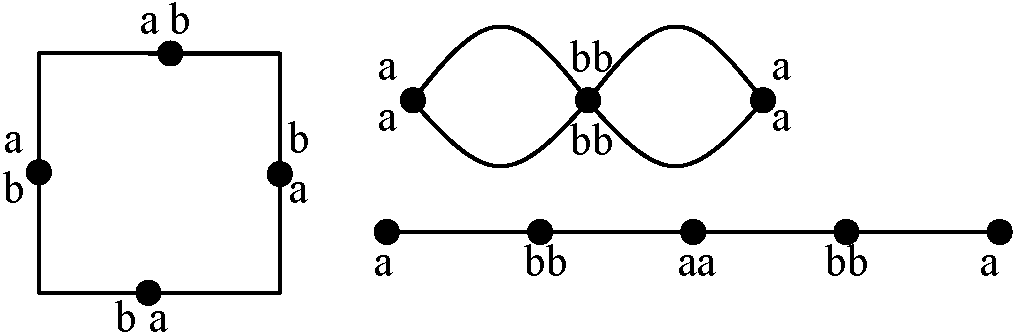
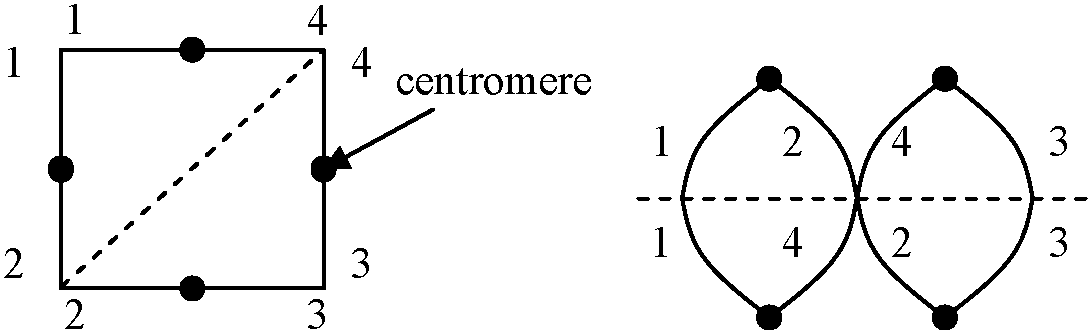
多価染色体: 3個以上の染色体の対合。対合のためには、相同的な位置(キアズマ)の存在が必要であり、以下に示すパターンが殆どとなる
染色体数・構造異なる親 → 染色体組合せに特徴的不均衡・特殊性持つ
(12, 14)/(23, 24): 1が存在しない = 不稔
→ この比率が花粉の稔性の出現率を決定している 逆位異型接合体 inversion heterozygosity_a__b__c__d_____e__R.D============8===== → 対合 ============8===== 遺伝 _a__c__b__d_____e__R.D 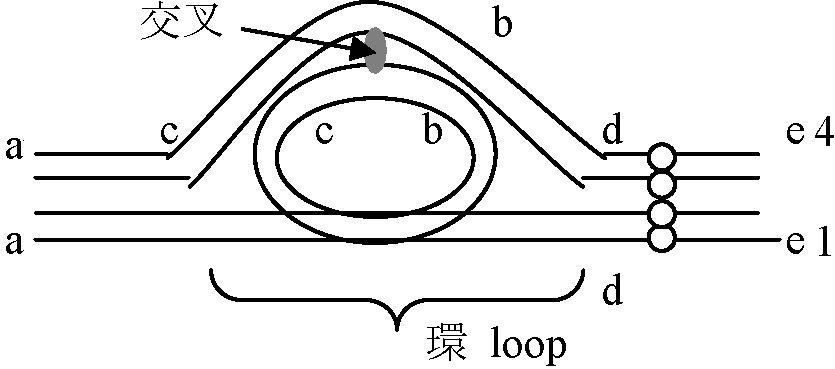 多糸期分析 pachytene analysis
ここで交叉の起こった時に4つのクロマチンの遺伝子配列パターン見る 2ヶ所に動原体があり重複有 → 染色橋chromosome bridgeの形成(動原体が2つあるため両極に引かれる)
2. a b c d ○ e 動原体がなく欠損している → 遅滞染色体 laggards 4. a b c d ○ e |
| 染色体観察 | |
|---|---|
|
染色体識別に最も良い時期は染色体がcompact構造をとり表れる分裂中期
体細胞分裂: 2n個染色体 ⇔ 減数分裂: n個の二価染色体が現れる 核型中期染色体観察 → 大きさ・動原体位置・二次狭窄や付随体の有無
→ 個々染色体識別 → 核型決定
種々要因に対する染色体の部分的反応差を利用し現れる分節構造により更に詳細な染色体識別も可能 染色体観察に適した材料
|
方法成長中根端: アセトアルコール固定、1 N塩酸60°C加水分解(時間は材料による)、フォイルゲン染色後、スライドグラス上で材料押潰し細胞散らし観察葯: 中身(花粉母細胞)をスライドグラス上に取出しアセトカーミン染色し観察 観察スケッチ: 染色体がよく分散し区別しやすい細胞を選び観察
材料: P. hexaphylla葯分裂細胞及びその前後段階の細胞。既製プレパラート 方法:
Ex. 化学物質が細胞分裂に与える影響 effects of 5-amino colchicine on cell division |
[ 生殖 ]
行動遺伝学 behavioral genetics行動特性の遺伝的基盤を研究 - 動物行動個体差の遺伝機構
⇒ 行動特性の起源や進化 ☛ 行動学 behavioral science (心理学) リスクは生活習慣・環境の方が大きく影響 研究法: 染色体異常(☛ ダウン症)、双子研究、養子研究染色体異常 Ex. レッシュ・ナイハン症候群 Lesch–Nyhan syndrome, LNS
自傷行為(2歳過ぎから観察される) + 他傷行為 痛風患者は男子に多いことに関係 1683 Sydeham, Thomas (1624-1689): "Treatise on gout and dropsy"
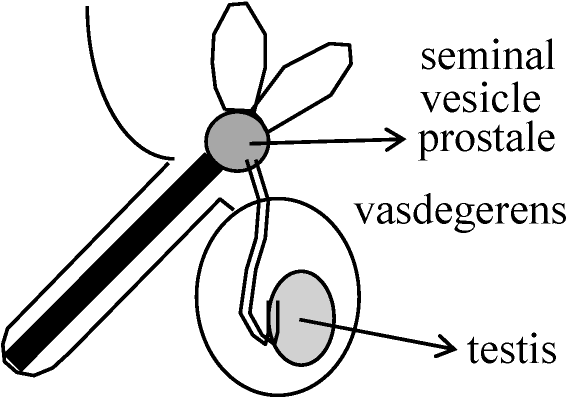
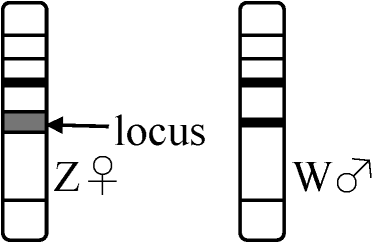
臨床観察 = 痛風が上流階層(過食過飲)に多 1983 西園: 躁病 - 尿酸値高い傾向 染色体細胞学 (chromosome cytology)性染色体 (sex chromosome): 性決定遺伝子存在 = (X, Y), (Z, W) ⇔常染色体 ≠ 性染色体 I: 雄ヘテロ: 雄が異なる性染色体を1本づつ持つ → 性染色体 X, Yと表わすP 細胞体 生殖細胞 F1 細胞 Ex. a. XY型 ♂ 2A + XX A + X ♀ 2A + XX ヒト、ショウジョウバエ b. XO型 ♂ 2A + X A + X ♀ 2A + XX バッタ、トンボ
c. ZW型 ♀ 2A + ZW A + Z ♀ 2A + ZW カイコ、ガ d. ZO型 ♀ 2A + Z
A + Z
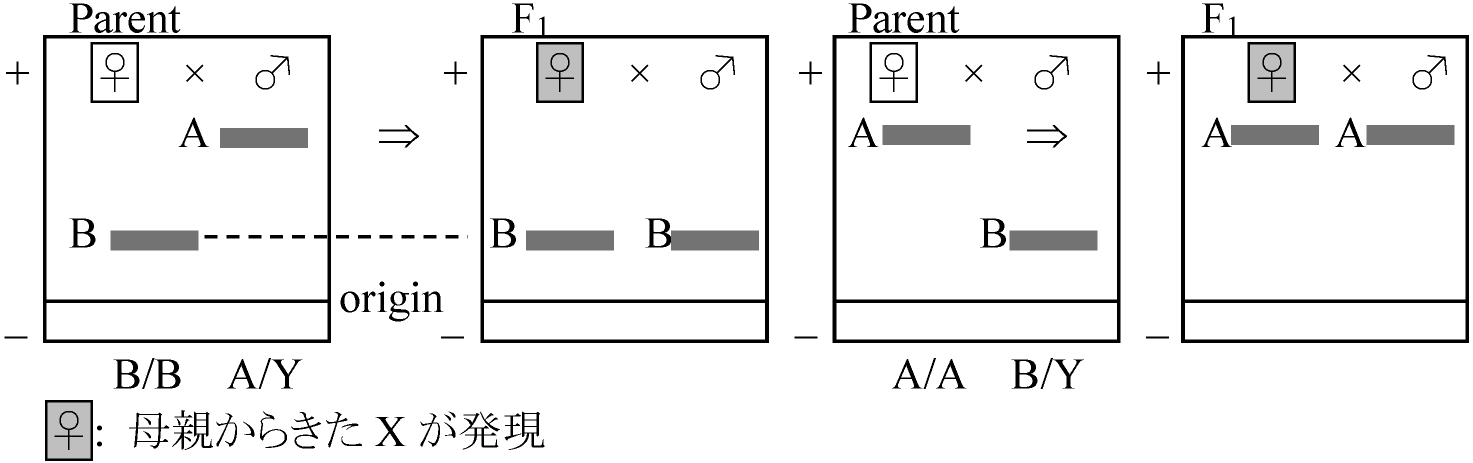
♀ 2A + Z スッポン、トカゲ、ハト 伴性遺伝 sex controlled (sex-linked) inheritance遺伝子が性染色体(X or Z)にある場合
X: ♂ = 多く表れる vs ♀ = 少なく表れる
♂ XY: XnY = 正常, XbY = 異常 Ex. ショウジョウバエ白眼 伴性遺伝は正逆交雑(相反交雑, 相反交配) reciprocal cross の結果異なる 相反雑種reciprocal hybrid 十文字遺伝: 伴性遺伝のF1で、父形質が娘に、母形質が息子に表れる限性遺伝 sex-limited inheritance遺伝子が性染色体(Y or W)にある場合限雄性遺伝 = XY型: ♂のみに表れる ⇔ 限雌性遺伝 = ZO型: ♀のみ Ex. グッピー(XY型)斑紋 – 雄のみに斑紋出るEx. カイコ(ZY型)体色 不完全限性遺伝: XY染色体上に遺伝子がある Ex. メダカの体色 従性遺伝 sex-conditioned inheritance遺伝子は常染色体上にあり、性ホルモンの働きで形質の優劣決まるEx. ヒトの若ハゲの遺伝
P
♀ × ♂ ♂: 禿る = A, 禿ない = a ♀: 禿ない = A, 禿る = a 母性効果 maternal effect
厳密には、キセニア・細胞質遺伝が非メンデル性遺伝 1. 非メンデル遺伝a. キセニア xenia: 父親の優性形質が直ちに母親の胚乳に表れる現象Ex. イネ(Oryza sativa)の形質遺伝(胚乳形質)
ウルチ = A, モチ = a 風媒花 = 比較的良く起こる Ex. トウモロコシ(Zea mays) 風媒花 - 砂糖性(ss)とデンプン性(SS) b. 細胞質遺伝 cytoplasmic inheritance: 細胞質(特に色素体・ミトコンドリア)が遺伝に関係 = 精子に細胞質殆どない → 卵細胞細胞質が形質決定 Ex. オシロイバナ葉色(全緑葉、斑入) c. 遅滞遺伝(母性遺伝): 一代遅れて形質発現 Ex. モノアラ貝の殻の巻き方: 右巻き dextral = L, 左巻き sinistral = l (メンデル遺伝する)
殻の捩れ(巻方)は卵割型で決まる = 卵段階(成熟分裂前)で決まる = 精子の入る前に決まる キヌレニン kinurenin (前駆物質) → black pigment: キヌレニン(有性遺伝子支配)がないと黒眼にならない
A-a
aa: 幼生larva段階: 眼色は黒 - 変態後: 黒色が消える
→ その部分はキヌレニン生産するが、それが拡散し卵内に蓄積される
"O" 致死遺伝子: ホモhomoはありえないはずだが、致死に至らずとも発生遅延、再生力欠如・鈍化が見られる。さらに睾丸形成不全が雄では見られる(雌では一応発生する) /"+" 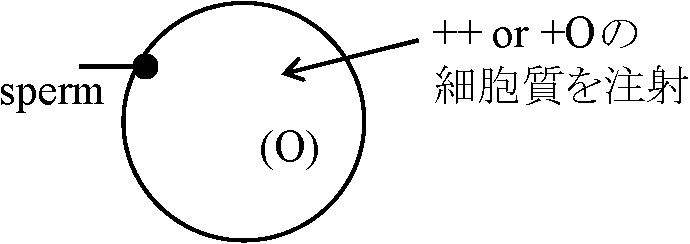
→ 殆ど全ての卵はgastrula段階まで発生できる
1) 3000 rpm → yolk↓ 1-3: 成分を別々に上澄と沈殿で注射すると全て上澄み注射時にgastrulaまで発生が進行する 4) 65000 rpm (1 hr) – ここで沈殿中にある – 分子量は大きくない = 可視的細胞構造物に結合していない
1-3上澄 a) 50-55°C処理、b) トリプシン処理 → 注射すると効果なし
「卵発生 oogenesis の雌による事後効果により特性決定がなされる」
→ 成熟卵にはm-RNAが残っている(卵においてはgastrula段階でm-RNA形成が開始される) 二型性 dimorphism性決定 sex determination, SDDef. 性(♂♀): 3定義存在1. 遺伝的性: 染色体構成による 2. 外観: 生殖腺分化の違い = 主に外性器、内性器
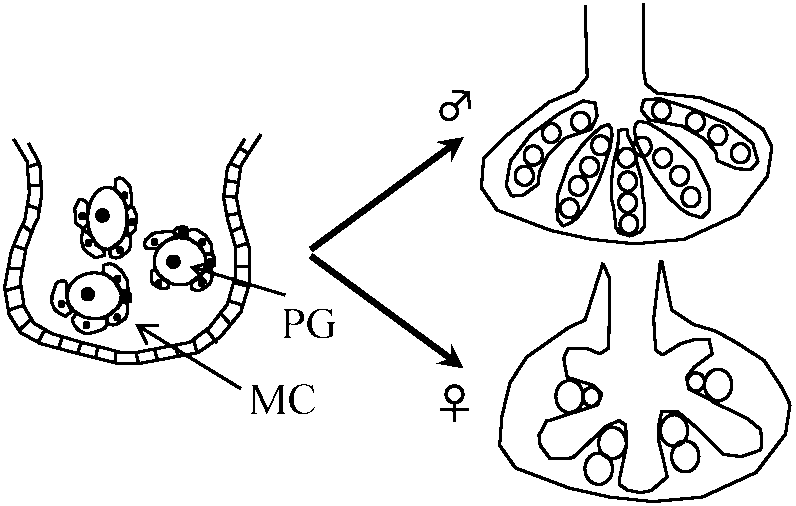
発生4 weeks (30 days): 性が肉眼で始めて確認できる時期 (gonadal ridgeは40 daysで確認出来る) 始原生殖細胞(primordial germ cell, PGC)の発生
付随生殖層変化、性分化発現理由不明 分化機構 differentiation mechanisms
♂ PGC → gonadal ridgeに入る |
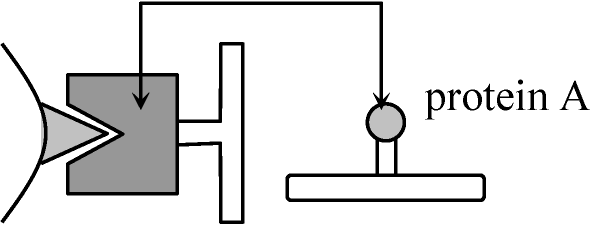
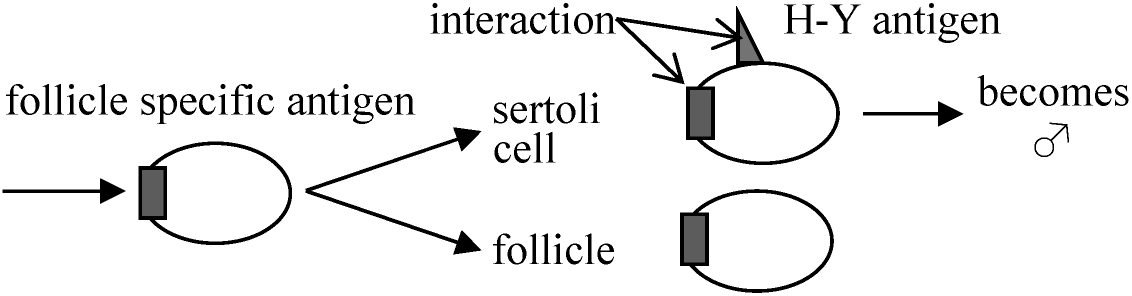
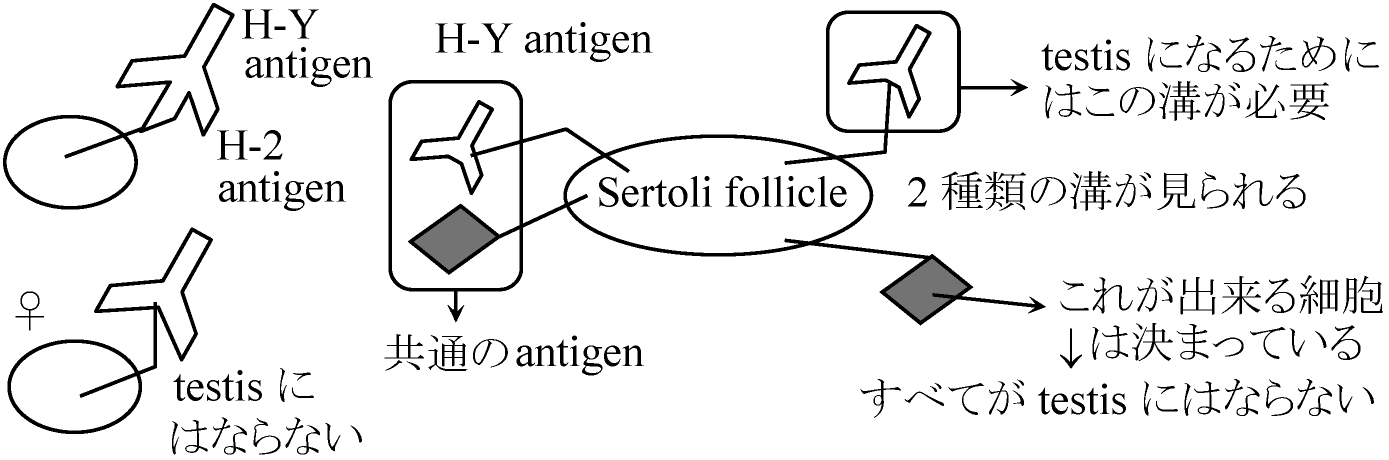
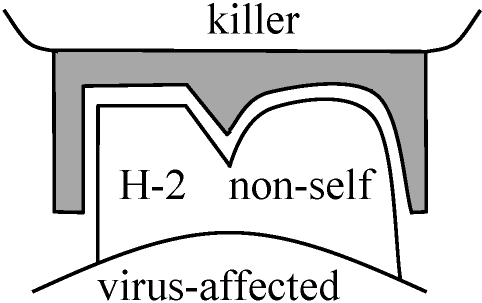
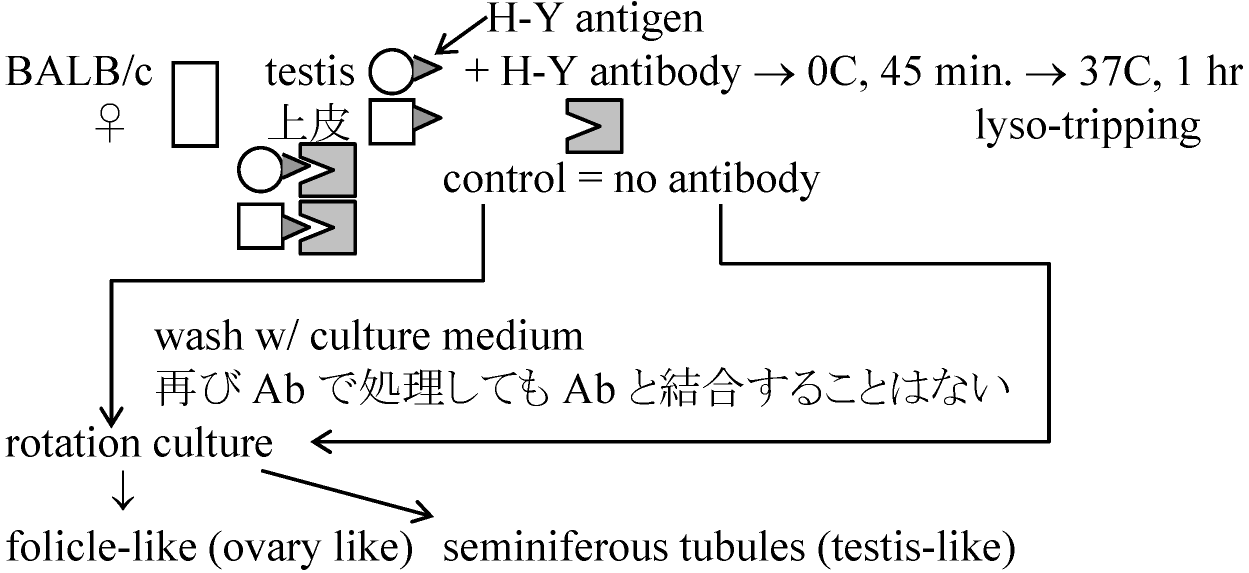
1978 Ohno et al.: H-Y antigen
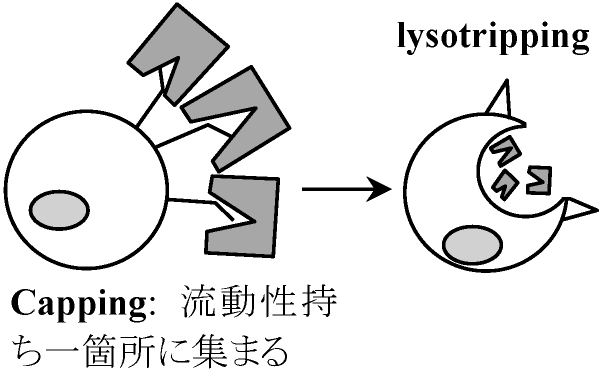
lyso-tripping: 昇華されてなくなる
H2N-
chimeric mouse 8 cell
→ BALB/c♀とchimeric mouseの♂をかける
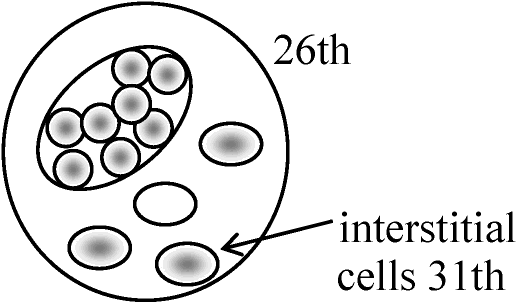
XX ⇒ protease (or sollagenase, protein分解酵素)で分解しsertoli cellだけを分離 生後3ヶ月以降にsertoli cell崩壊interstitial cells増加 → これをproteaseで分解しinterstitial cell(H-Yを持つ)を分離 ① H-2d/H2d♀にH-2dのsertoli cell注入し抗体(antibody, 抗体①)を作らせるcytotoxic primordial sertoli I.C. follicle lutein ♂ ♀ testis germ cell cell cell cell 1) Unabsorbed Ab (①, ②) - +++a + ++ - +++b - 2) ♂は脾臓♀はモツと共に - +++c - ++ - - - abosrbed H-Y antigenで吸収する 3) Absorbed with follice cells - - - - - - -
+++: 非常に強い反応, ++: 強い反応, +: 反応がある, -: 反応無し
感作させた♀ → 脾臓取り出しT-cell分離 ♀ T-cell H-2 type serotoli ♂脾臓 ♀脾臓 ♀ follicle cells H-2b H-2d/H-2b H-2d +++ (H-Y)1 · · · H-2k - H-2d H-2d H-2d ++++(H-Y)2 +(H-Y)2.1 - H-2k ±(no H-Y) - n.d. H-2k H-2k H-2d +++3 - · + H-2k +++ - n.d. H-2b + n.d. n.d. H-2d H-2d H-2d ++4 + - · H-2k ++ - n.d. H-2d/H-2k n.d. + n.d.
n.d.: no data available
H-YよりH-Yでないものが大きく作用していると考えられる H-2 unrestricted (H-2 typeには関係しない反応) sertoli-follice specific reaction → angiten存在 ☆ 脾臓に関してはH-2 restricted(限られた)反応Sertoli cellに関してはH-2 unrestrictedな反応 ⇒ Sertoli-follice specific antigenの存在(この際はH-2 unrestricted): H-Yに関する限りH-2 restricted
Y-chromosome (Homo sapiens研究から) 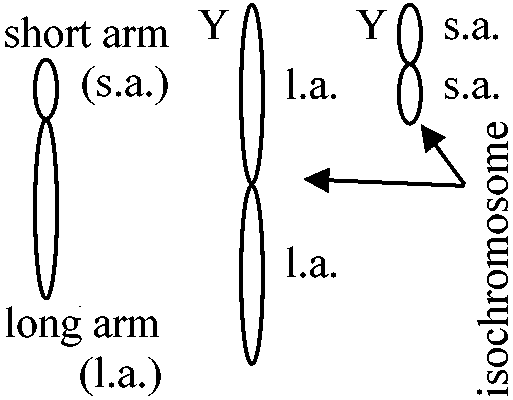
isochromosome.のH-Y antigen white leghorn = loss brown - sex linked: feather color ♀brown/♂white
12日目以降はfeather colorでの♂♀区別不可能
ZZ: ovary antigen
spleen(脾臓) ♂ → ♀antiserum: normal ovary/estradiod-treated ovary(=ovotestis) 1, 2の皮膚細胞に対する毒性
にもH-YがありH-Yの産生にWは関与しない
unabsorbed ⇔ +++(H-Y present)
sex-revsersed ZW♂ × WW(♀) → ZZ(♂): ZW(♀): WW(♀) → WにH-Yのstructural geneはない ZZでもH-Yができる ZZ testis cell (Xenopus) + H-Y antigen (mouse)
毒性消失 → H-Yはtestis cellに付く(生殖巣のみ、体細胞につかない)
testisの中にはtestisの分化を促すような物質があるのでは? 性決定に関する他の条件1947 Jost: ウサギ rabbit
♂胎児去勢 → Müllerian duct残る(普通はMüllerian hormoneのために消える) → 輸卵管・子宮が出来る
ヒト♂ pseudohermaphrodite → Wolffian duct(男性化の兆候無し), urog.sinus, 外陰部 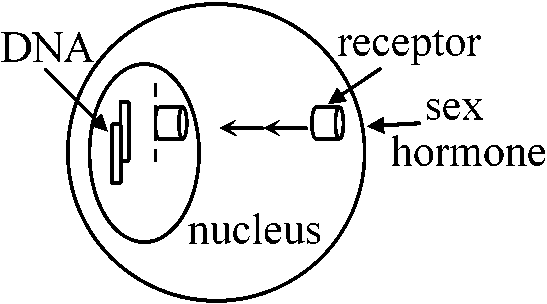
receptor: 特定DNAに働き活性高める → sex hormoneにはreceptor必要 TESTO 4 ng/mg in testis
84 d___5 ng/mg (peak) → Wolffian d.の雄化盛ん(U.S.) TESTO synthesis → 母体のchorinonic gonadtropin [胎盤ホルモンによって推進される] 半分抑制された状態 ∴ もっとTESTO投与すればmale化する
androgen
choresterol → pregneolone → progesterone → androsterodione → T → estradiol
表. ヒトの性染色体 (染色体数, 性染色体構成) Ex. 47, XXY = 47本, XXY)
1938 Turner: ターナー症候群初報告 - 生殖腺無形成症、生殖腺発育不全症数, 性染色体 名称 特徴(事例 → 皆がそうではない)
1/3000: 外性器女性 - 二次性徴発現なし
1/1000人: 女性化乳房、矮小睾丸、髭・腋下・恥毛発育不良、声
精神障害(精神分裂様症状、精神遅滞)に関与
超女性 superfemale (誤解を与えるため非推奨呼称) グエベドセス guevedoces (gneve = penis, doces = 12): 遺伝病 出生時に女子と思われる - 輸精管・前立腺存在(必ずしも正常ではない)
精巣 testis (精子形成 spermatogenesis): 正常 E.G. (external genitalia) → 雌型: serotum → labia-like, u.s. (urogenital sinus 尿生殖洞) → blind vaginal pouch, clitolis → like phallus
突然: 筋肉発達、声帯発達(男声)、penis発達、scrotum(陰嚢)発達 一方、前立腺小さい、ヒゲ生えない ⇒ T.M.F. (typical male form)は精神的にも女性化するがguevedocesでは全く男性化
13家族24人 = pseudohermaphordite
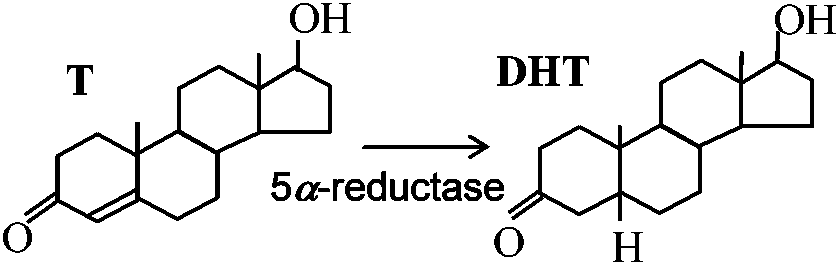
5α-reductaseがあれば男性化するがgneve docでは量が少ない 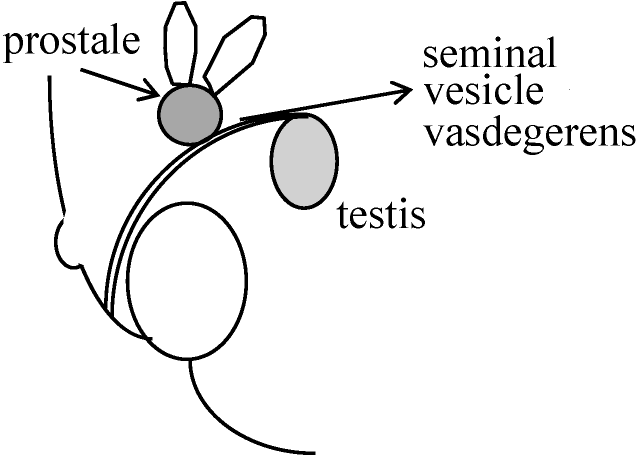
① 少ないながらもある程度DHTができるから正常に近くなる XY, XX = 必ずXがある X-chromosome: 生体にとって有利 → ♀に有利 ↔ 2つあることによって不利になる = ♂に有利 1) 性染色質 sex chromatin 1949 Barr & Bertran: ネコ♀神経細胞
1949 Barr & Bertran: ネコ♀神経細胞
核小体: ♀のこれに瘤が高頻度 ↔ ♂にない 2) 性染色質との関係
number of sex chromatin
2N = 46 = XX + 2A
Ex. 2N = 2 - 1 = 1, 4N = 4 - 2 = 2 3) Late replication of X chromosome1960 Taylor: Chinese hamsterの培養細胞
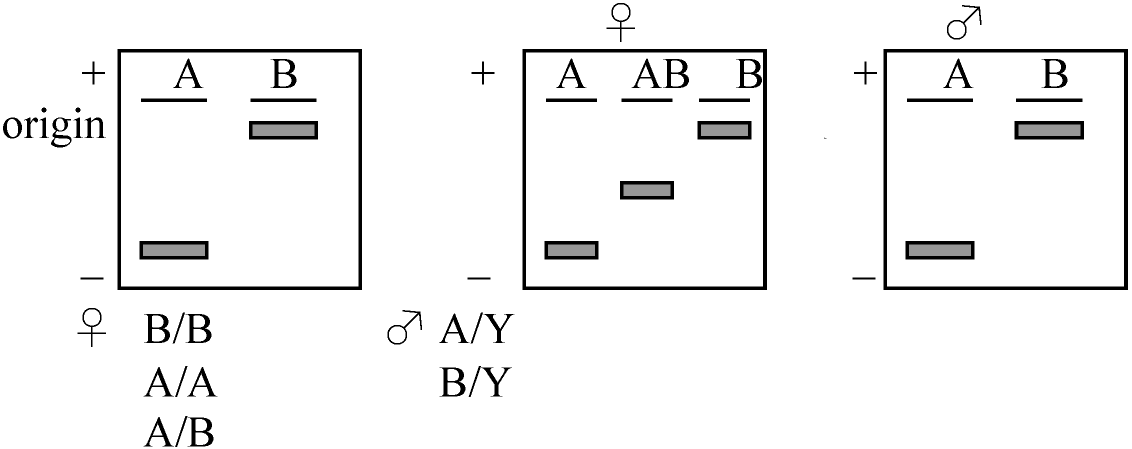
同調培養(全細胞が同じ分裂期にあるよう調節) 4) Sex chromatin is on X (コブはX chromosomeである)1959 Ohno: rat liver5) Single active X theory1961 Lyon: 仮説 sex chromatinがX-chromosomeであれば、それは不活性なX chromosomeであろうCattanach translocation-cのgeneに注目
P/P - pink eyed, +/P - black, +/P/P - black (blackになるはずだが実際はvarigationになる)
例外: 生殖細胞__inactivated ×
[wild = B] vs [mutant = A → single amino acid substitution グルタミン → グルタミン酸]
mutant polypeptide A
B wild peptide
もし活性が同じ(同じ速度でpeptideを作っていたとすれば) → AA:AB:BB=1:2:1できる
heterozygous♀(A/B)皮膚細胞 → 組織培養(維芽細胞が一番増殖容易)
Expected = クローン増やしてもAB出てこない
(1) Hypoxanthine-guanine phosphoribosyl transferase (HGPRT) Nitro BT (人工色素, pale yellow, soluble) + NADPH + H+ → formazan (色素還元型, blue, insoluble pigment) ⇔ この説で説明できない事例: the inactivation occurs at random (2)
カンガルーについて良く調べられている
__________________________________ 2. sex chromatin = on X 3. X → DNA late replication 4. single active x theory genetical inertness of a single X(遺伝的に活性を持たないX) 3 = 4のつながり
horse(♀) × donkey(♂) → mule これを用いた G6PD導入によりs.c.が遺伝的に活性を持たないXであること判明 Xが2本とも発現しないのは何故?
XXとXY: 2本とも働いたら不公平が生ずる。物質を多く合成することで自然浮動が動くと困る |
|
遺伝子/染色体変化 = 遺伝 ↔ 個体変異: 一時的 (× 遺伝 ≠ 遺伝子関係) 1) 染色体突然変異 a) 染色体数変化 Ex. 倍数性・異数性 b) 染色体の部分的変化: 染色体の一部が …
欠失: 消失 Ex. 猫啼病: 第5染色体短腕の一部消失 相互転座: 相同ではない2つの染色体間での部分的交換 重複: 複数回繰り返される - 生存に影響小
新たな遺伝子形成の可能性 - 進化的に重要 逆位 inversion: 逆向きにつながる 2) 遺伝子突然変異: DNA塩基配列変化 → (1)よりミクロなレベルで変異a) 点突然変異 point mutation 非同義置換 non-synonymoussubstitution
ミスセンス突然変異 missense mutation: コドン変化によりアミノ酸置換 サイレント変異 silent mutation: アミノ酸変異なし b) フレームシフト frame shift
挿入 insertion: 塩基配列加わり変異 Ex. ミスコピー → 大部分は修復 Hb: ヘモグロビン 鎌形赤血球貧血症 sickle cell anemiaアフリカ(特に中央アフリカ、マダガスカル島) - 遺伝するHbはO2失う → 溶解度低下し赤血球中に細胞破壊する結晶物質作る → 遊離した水は赤血球外に出て鎌形化 → ガス交換率低下 + (赤)血球は粘性増し毛細血管を塞ぐ → 血管障害
ホモ = 2歳未満に殆ど死亡 1949 Pauling, Linus Carl: Hbタンパク質部分の差異が原因 → 分子症 1956 Ingram, Vernon Martin: Hb N末端から6番目アミノ酸変化 = Glu → Val
アミノ酸だた1つの変化 → 個体の生死に関わる変化
βグロビン遺伝子6番目コドン変化: GAG (Glu) → GTG (Val) |
ヘモグロビンβ-鎖一次構造
S: sicle鎌
チロシナーゼ(酸化酵素)の働きが先天的に不完全 → メラニン全く(か殆ど)合成されない
放射線 (Ex. X線)、紫外線、活性酵素、化学物質 (≈ 発癌物質)、トランスポゾン 突然変異mutationの推定グルタミン酸code: GAA, GAG → バリンcode: GUU, GUC, GUA, GUG
突然変異は同時に2個所起こるのは稀 → 1個所のみ変化したと仮定
6番目のアミノ酸がGlu → Lysとなったもの ハプロタイプ (haplotype)同じ染色体のある領域に並ぶマーカー(SNP)の組合せのセットHapMapヒトゲノムにおけるDNA多型(SNP)のパターン(ハプロタイプ)地図 (国際ハプロタイプ地図作成プロジェクト作)ハプロタイプネットワーク (haplotype network)各ハプロタイプ間関係を示す - 塩基配列近いハプロタイプを同色で示すこと多SNP (single nucleotide polymorphism), スニップ一塩基多型。一つの塩基が他の塩基に置き換わっている多型
ヒトゲノムでは約1000万箇所以上のSNPを確認 → |
|
対合型異常対合: 10II (bivalent) + 10I (univalent) 一価の分配異常: 10Iが任意に分配不稔: 一価の分配異常のため 退行現象 restitution: laggardsが第一分裂終期あたりまで残ると隔壁できない この膜は稔性有 → 子孫残す可能性 連鎖(連関、リンケージ) linkage= 2つの遺伝子形質を表す遺伝子が同一染色体にある場合Ex. (AとB), (aとb)が同一染色体上にある → (AとB, aとb)は連鎖している 連鎖遺伝子 linkage genes連鎖群 linkage groups: 一緒に遺伝する形質の集団 連鎖群数と染色体数(n)は大部分の生物に関しては一致する
例外 連鎖群数 染色体数
⇐ 原因不明: 不活性遺伝子(染色体)の存在、異質染色体との関連等が考えられているが、実験的証明なし
⇓ P: AABB (AB/AB) × aabb(ab/ab)
|♀/♂ AB ab
交鎖率 (%) = (交鎖により生じた生殖細胞数)/(全ての生殖細胞数) × 100 F1 (AB/ab) × [劣性ホモ] (ab/ab) → 交鎖率(%) = (交鎖により生じた子数)/(検定交配での全子数) × 100 連鎖と組み替え BL (m):Bl (n):bL (n):bl (m)
mBL nBl nbL mbl 同じ連鎖群に含まれる形質(A, B, C)に着目し交配実験 |
原則: 遺伝子は染色体上に線上に配列
交叉はランダムで、その頻度は遺伝子間距離に比例
B 9% A 6% C
染色体は切れやすい所と切れにくい所(染色体の太さや括れ)が存在し全く同じではない 染色体数変異染色体数 (Sinoto 1926)。染色体組 chromosome complement (set)表現法 Rice Corn Wheat Human 生殖細胞の染色体数(単相世代) n 12 10 21 23 栄養体・胞子体(複相世代) 2n 24 20 42 46 基本数 x x=12 x=10 x=7 x=23 原始基本数 ↑ proto-basic numbers b b=6 b=5 b=6 9 b=8 basi-basic numbers b=3 ↑ 減数 原始基本数は説であり断定的なものではない 染色体異常構造変化を伴う異常 structural aberration構造変化のない異常 non-structural aberration (粘着性 stickiness によるものあり) 染色体の構造変化 structural changes in chromosomesゲノム genomeWinkler (1920, 当時の定義): 一組の半数染色体木原: 生物生存上不可欠な染色体一組 現在: ゲノム = DNAの総体 = ある生物が持つ遺伝情報全体
真核生物: ゲノムは核、ミトコンドリア、葉緑体に独立のゲノムを定義
ゲノムを反映しているはず |
|
1916 Winkler H: 近縁種、品種等の間で染色対数に増減の見られる現象 Def. 新倍数体 neopolyploid: 新たに形成された倍数体 真数性 (正倍数性, s.l.) euploidy基本数の完全な整数倍
コムギ: 2n = 14 (2x), 28 (4x), 42 (6x) 異型倍数性 heteroploidyEx. Artemisiaの染色体数
フクドヨモギ
イヌヨモギ
オトコヨモギ
ヨモギ 倍数性 (正倍数性, s.s.) haploidy基本数basic number (x)の整数倍の染色体組(ゲノム)持つ= 倍数性はゲノム重複現象 (一般に2倍体(2n)が基準) Ex. 1倍体(半数体) haploid, 2倍体 diploid, 3倍体 triploid, 4倍体 tetraploid, 5倍体 pentaploid, 6倍体 hexaploid, 8倍体 octoploid … 動物では稀(正常発生稀) ↔ 植物では普通Def. 混数性 (体細胞倍数性): 1個体内の組織・細胞間に倍数関係がある場合 異数性 aneuploidy (inclucing monosomics and trisomics, s.l.)染色体の一部増減による不完全な時 - 近縁種間自然倍数種の多くは異数性
高次異数性 hyperaneuploidy: 正常より染色体数が多い Ex .1. ダウン症候群(蒙古症) Down syndrome (1866 Down 初記載)
2n = 47 (∵ 21番染色体3個 = 3数性 trisomy)
母親 25(歳) 1/1400 30 1/1000 35 1/350 40 1/100 45 1/30
症状(※ 必ずではない): 発達遅延、身体的特徴、知的障害 Ex .3. ヨメナ: 2n = 51, 52, 53, …, 76 Ex .4. Paphinopedilum sp.: 2n = 26, 27, …, 44 Ex .5. Lycoris 2n = 22, …, 2n = 33, …, 36 (日本産33のみ → clone増殖) 他例: シライトソウ、ショウジョウバカマ、アオミドロ
尾瀬 2n = 26, 30, 31, 32
polyploidyの典型と考えたが、染色体数増すにつれ大きな染色体減る = 偽倍数性 agmatoploidy 同質倍数体 autopolyploid, autoploidy同質4倍体 autotetraploid: 同一種由来のゲノムを4有する同質3倍体 autotriploid: 同一種由来のゲノムを3有する 異質倍数体 allopolyploid, alloploidy異質4倍体 allotetraploid (複2倍体 amphidiploid)ゲノム解析 genome analysis染色体組の各構成染色体の遺伝的相同性(遺伝子分析) 1918 坂村徹 (1888-1980): コムギ属倍数性発見 x = 7Ex. 木原: コムギ (Triticum L.). 芳賀・倉林: 日本産エンレイソウ (Trillium L.)
未知 (Case. 異数性分析Ex. ホウレンソウ
2n = 12 (x = 6), or 2n = 13 = 12 + 1 → 6組の染色体組合わせ 倍数体の優位性 (superiority of polyploid)自然界では一般に倍数体のほうが生存に有利 (安定化した所で倍数化起こりやすい)。人為的倍数体は劣勢であり、倍数化の筋道が自然なものと異なる
安定化: 多価染色体減少 = 2価染色体増加 (2価染色体形成遺伝子の存在あるいは新しく形成) 1914-1915 田原正人(1884-1969): キク属染色体数は9の倍数 - 倍数性発見
シュンギク 2n = 18, フランスギク 36, コハマギク 90
2x: 雑種性 – AB → 受粉 restitution → AABB × ♂ ノヂギク 2n = 54 (6x) JJJJJJ |
F1 2n = 63 MMMMJJJ こちらだけ倍数化
倍数体の安定性獲得 diploidization 個体変異の原因1) 2倍種間の分化
P_______________________________F1 (2n = 18) → 複製点数同じ: 大きさの変化が種分化に関与 2) 2倍種内の分化
アブラギク__北九州_近畿___関東
______ リュウノウギク: 染色体分化見られない + 連続分布
ワカサハマギク 2n = 36, 4x: MMBB ecotype 多型 →----┐ キク科植物のgenomeは同質で対合パターンが多い
MM MM: x = 9
約190万の組み合わせが考えられる 遠縁交雑 wide crossing属間雑種多属間雑種(最大6属間雑種既知) → 形態変化大きい種分化はそれほどない? 機構
Ansellia, Maxillaria, Stanlopea, Calanthe, Zygopetallum 広く分布、砂漠植物 2n = 4発見 – 現在知られる最小染色体数。発芽試験を行うと稔性ない = H. gracilis KH-1 (K: Kanthus, H: Hiroshima. 広島大学の株) 染色体異常高率発生だが、1年生植物なため子孫にその遺伝子伝わりにくい。生活環約9ヶ月。クローンの異常染色体保持を行うため栄養繁殖を試みると、挿木は死滅、カルスは苗化困難だった。カルス形成過程で緑色スポット(苗状原基)得られ、これは増殖率が一週間あたり4nと高く、これを用い研究行う 染色体の安定性: 染色体により異なる。安定な染色体と不安定な染色体を組み合わせると不安定な染色体も安定化する → 育種、細胞分裂および細胞融合の研究に適しているなど応用可能
プロトプラストから細胞融合 - 他種に染色体埋め込み苗化試験(クレピス)
染色分体 chromatid |
[ 進化論 ]
|
対立遺伝子頻度gene grequency変化を研究する遺伝学の1分野
目的: 集団における遺伝的不均一性と関連する遺伝構造の解明 || Def. 遺伝子頻度: 集団内で対立遺伝子alleleが含まれる頻度(割合)Ex. A A A A a a a a a a → A遺伝子頻度 = 4/10 突然変異 + 自然淘汰 → 遺伝子頻度変化 → 進化機構解明 || Def. 遺伝子型頻度: 集団内である遺伝子型genotypeの頻度|| Def. 表現型頻度: 集団内である表現型phenotypeの頻度 Ex. AA AA Aa Aa aa → AA遺伝子型頻度 = 2/5, A表現型頻度 = 3/5 メンデル集団 (Mendelian population)Def. 遺伝子プール(給源) gene pool: 遺伝子の集合として交配可能な集団Ex. 個体数 = N → 遺伝子プール = 2N
→ 時間・空間共に連続で、ある個体群に属する生物
実質的遺伝子交換のある集団 → 個体群動態 demography = 動態統計的研究 (Pianka 1978)
1. 集団が十分に大きい
2. 交配は自由randomに行われる 「メンデル集団内は遺伝子頻度も遺伝型割合も、世代が経っても変化しない」 Pr. A:a = p:qの比で存在 (p + q = 1)
任意交配の結果は (pA + qa)2 = p2AA + 2pqAa + q2aa aの頻度 = q2 + 1/2·2pq = q(p + q) = q
∴ A:a = p:q
⇒ p2AA:pqAa q2aa = const. // (p1A1 + p2A2 + p3A3 + …)2 = p12A1A2 + … 1) 自由交配(任意交配): 生活力に差のない形質を考え自由交配を続ける →集団内遺伝子頻度は一定を保つ (≡ ハーディ・ワインベルグの法則) 2) 自家受粉: 続ける → 遺伝子頻度 = 不変
ヘテロ↓ ホモ↑ (ヘテロ%, F1 = 100%, F2 = 50%, F3 = 25%) 近交係数, F Ex. 自家受粉 F = 1/2 同類交配 assortative mating: 表現型が似た個体間でより交配が起こる 遺伝的平衡Ex. 軟骨異栄養症 chondrodystrophia: 優性突然変異遺伝子によるEq. Haldane, John Burdon Sanderson (1892-1964): 2v = x(1 - w) ⇒ 平衡状態
v: 突然変異率 (軟骨異栄養症が毎代出現する率) (♂♀側があるので2倍) x(1 - w): 毎代に集団中から患者が除去される量 Ex. if x = 1/9400, w = 1/5 (デンマーク) → v ≈ 4.3 × 10-5 近縁係数: メンデル集団であれば
Ex. 兄弟 = 1/2 (半数生物および伴性遺伝の場合は異なる), いとこ=1/8 中立説 neutral theory1968 Kimura M (木村資生, 1924-1994): 中立説Def. 遺伝的浮動 (random) genetic drift: 偶然発生した変異(遺伝子) → 固定 1969 King & Jukes 1969: 非ダーウィン進化 non-Darwinian evolution 中立説指示: 分子進化での突然変異遺伝子の種内蓄積は自然淘汰よりも遺伝的浮動による偶然的固定の結果 Def. 機械的浮動 Ex. 壜首効果 bottleneck effect進化速度 (単位: 年、世代): k = f0vT f0: 突然変異のうち自然淘汰に中立なものの割合
機能的制約大 → k = 小 (Ex. 機能的制約 = 0 → k = 1
vT: 総突然変異率 遺伝子淘汰 genic selection: 淘汰が直接、遺伝子に働く場合 Ex. A: 正常(野生型)遺伝子, A': (淘汰上有利な)突然変異対立遺伝子 A淘汰値 := 1 ⇒ A'淘汰値 := 1 + s (s ≡ Aの有利さを表す淘汰係数)
淘汰値: AA = 1. AA' = (1 + 2s) A'A' = (1 + 2s)
ln(pt/qt) = ln(p0/q0) + st (q0 = 1 - p0) pt ≥ 99.9に係る世代 → 13813.5 (1世代1年 → ≈ 1400年) 明らかに有利な突然変異遺伝子は短時間で野生型と置換 淘汰方向
密度依存性density-dependent淘汰: 遺伝子型適応度が集団密度で変化 頻度依存性frequency-dedendent淘汰: 遺伝子型適応度が頻度で変化 Ex. 少数者有利 majority advantage: 低頻度 - 有利 ⇔ 高頻度 - 不利 平衡多型に関与(稀)
安定化淘汰 stabilizing selection: 極端な個体除去する淘汰 - 現状維持的役割 ⇒ 中央の量的形質残る
= 正常化淘汰 normalizing selection, 求心性淘汰 centripetal selection |
体重が極端に重いものも軽いものも中間のもにに比べて高い 方向(指向/定向)性淘汰 directional selection: 量的形質平均値が最適値とは違った位置にある場合に起こる ⇒ 最適値に向かい進化環境条件が一定方向へ変化している時に見られる 分断(分断化/分断性)淘汰 disruptive selection: 1集団に対し毎代最適値が2つ以上ある = 最適niche複数あると起こる分断淘汰 ≠ 集団中に2種類の対立遺伝子 → 淘汰方向が世代毎に逆転 全淘汰指数 index of total selection (Crow 1958): 淘汰の可能性(上限値)= 子数の(分散)/(平均)2 堅い淘汰 hard selectionと柔らかい淘汰 soft selection___________________Hard selection________Soft selection___________________C = constant_________C = flexible 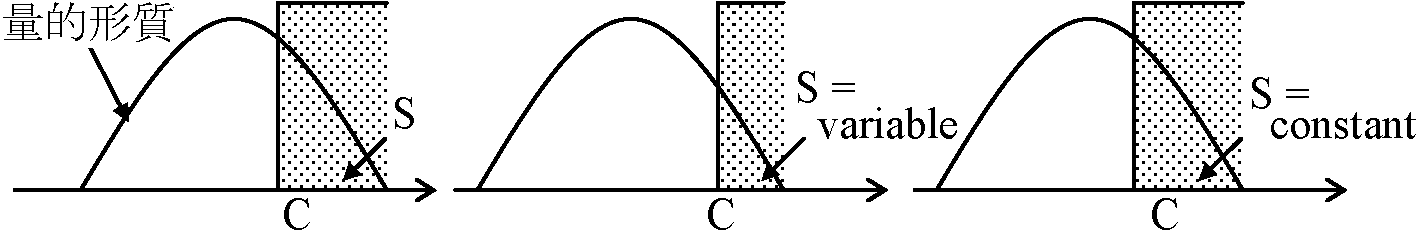 ___________________→ 量的形質が下方へ変化 堅い淘汰: 淘汰により残る形質数減少 ↔ 軟らかい淘汰: 変化しない 集団内変異(個体群内変異)多くは正規分布遺伝的なもの + 環境的なもの → 選択selectionを用い確認可能
Ex. 人間の背丈: polygenes支配 (集団内における)多形一般にtransitional - 周りの環境による平衡多型異型接合型が最も生存力が強い → 多面発現効果 (pleitropic effect) Ex. オオシモフリエダシャク 平衡多型現象 balanced polymorphismDef. 遺伝的多型: 遺伝子頻度にし約1%以上の変異存在超優性 ≡ ヘテロ接合体が全ホモ接合体よりも自然淘汰に有利な場合 Def. 平衡多型: 超優性 → 2個の対立遺伝子が環境変動がない限り安定共存Ex. 近縁な2種のカタツムリ: Cepaea nemoralis, C. hortensis: 殻色
黄/淡紅/褐 + その上に0-5本の暗色の帯模様つく → 表. 芝地にまいた人工餌の7日間ずつの採食様式。*: 期待値より有意に多
ツグミ ムクドリ イワヒバリ スズメ
1) 実験1は180緑:20褐、2は20緑:180褐
→ 鳥は視覚で餌選択 → スズメでは頻度の高い方がより多く食べられる
近縁な同所的種はそれに似ていない表現型有する方が有利 (Grant 1958) 淘汰圧 selective force組み換えシステム開放系 open habitat → [spectrum] → 閉鎖系 closed communities実生seedlingから成熟matureまでの死亡率高い Ex. Fagus grandifolia: 1.0個体/10 years/0.1 acerの割合で実生生存可 淘汰圧: 閉鎖系に発達する組換システムとし開放型は将来の変化のため必要1975 Levin, 1976, 1978 Solbrig 閉鎖型といっても、その環境biotic environment (Ex. 病原菌, 草食動物)は一定ではない - それに対処するために開放型を有する 分子進化 molecular evolution分子進化速度Ex. ヒト vs ウマ: Hb-α鎖 141アミノ酸座位 - 18カ所で異なる
ヒト-ウマ分化(化石による推定) - 8000万年前
10億年に0.8個の変化 (ヒト・ウマ進化と2経路 → 2で割る)
N: 集団内個体数 (集団の有効な大きさ, Ne < N) Case. u = 2sNe/N ⇒ k = 4Nesv 分子(進化)時計 molecular (evolutionary) clock分子進化速度は一定 (ただし、遺伝子が異なれば速度も異なる)= 突然変異蓄積速度一定 → 化石資料なしに系統樹が作れる 保守性: 機能的に制約少ない(重要でない)遺伝子ほど進化速度大Ex. プロインシュリン: A(30アミノ酸), C(21), B(33)
Cが切り離されA-Bとなりインシュリンとなる(Cは不要)
同義的塩基置換: 頻発
vT: 総突然変異率 遺伝的多型 genetic polymorphism: 中立説 vs 平衡淘汰説染色体多型: 逆位 → 組み換え起こらない = 超遺伝子 supergene平衡淘汰 balancing selection(相反する方向に働く淘汰)により維持 + 頻度依存性淘汰 DNA多型 - 中立説を支持 Ex. インフルエンザウィルス、エイズウィルス |
|
生物適応の遺伝的基礎を探る = 生態と適応の遺伝学 (酒井 1976)
集団遺伝学: 遺伝子頻度変化機構の解明
↕ 橋渡し = 生態遺伝学
進化生態学: 自然選択・適応 工業暗化 industrial melanism産業革命: トン石炭煤煙/km2/月 (50トンに達したことも)樹木: 異常落葉、樹皮黒化 1848 黒色型(暗化型) Biston betularia がマンチェスターから初報告ついでヨークシャー工業地帯とその周辺から報告 1896 Tutt, James William (1858-1911, 英), 昆虫学: 鳥捕食仮説暗化型: 黒い幹の上でカモフラージュとなる (+ 中間型は稀) 1914 Bowater W: 中間型稀 → メンデル遺伝と推定1958 Kettlewell: 工業地区 = 9割が黒色型(1956年調査)
70種以上のガ(蛾)で暗化が工業地帯で見られる 土中重金属と植物1966 Jain & Bradshaw: Agrostis tenuisの耐銅性を水耕法で調べる→ 鉱鉱坑知覚のA .tenuisは耐銅性が高い 1967 Antonovics et al.: 高耐銅性植物は銅がある方が成長がよい↔ 低耐銅性植物は銅があると成長阻害 1968 McNeilly & Bradshaw: 銅汚染地区と非汚染地区
→ A. tenuisの個体と種子を採取
銅汚染地区 > 15 m → 耐銅性なし 採取種子から育てた実生と成草の耐銅性には高い相関(r = +0.97)あり、また実生の平均耐銅性は親よりも高い → 成体になるまでに耐銅性の高すぎる個体が淘汰される → 安定化淘汰 stabilizing selection 1974 Walley et al.重金属耐性個体は正常集団中にも少数ながら存在し、汚染土によるスクリーニングを受けた結果、耐性個体が残る |
1974 Gartside & McNeilly: A. tenuis, Lolium perenne, Cynosurus cristata, Dactylis glomerata, Poa tricialis, Arrhenatherum elatius, Anthoxanthum odoratum, Plantago lanceolata, Trifolium repensの耐銅性調査
A. odoratum, P. lanceolata, T. repens: 銅土壌で全枯死 → 耐銅性なし 稔性変化1973 Lefébvre: Armeria maritimaの自然状態における自殖率を調査 – 2型
A型: ヘテロ AC/ac 花粉網目、柱頭平滑 → 自家不和合性ありA-Bのみで交配可能
仮定: if 他殖率100% → A:B = 50:50, 自殖率増加につれB型が増す
鉛、亜鉛の耐性型と正常型(耐性なし)集め、それぞれの単植区と混植区を設定し競争試験 単植 混植 ホソムギ混植 型 耐性 正常 (繁殖比) 耐性 正常 (繁殖比) 耐性 正常 (繁殖比) ヌカボ 477 630 (0.757) 354 760 (0.465) 144 309 (0.466) ヘラオオバコ 791 1050 (0.753) 719 1230 (0.585) 794 879 (0.903) ハルガヤ 340 429 (0.793) 143 307 (0.467) 113 196 (0.576) 耐性型は、競争に非常に弱い – 競争条件下(正常地)では生存しにくい 煤煙による土壌汚染1975 Wu et al.
精錬所周辺9ヶ所及び正常地2ヶ所から土壌とAgrostis stolonifera採取 |

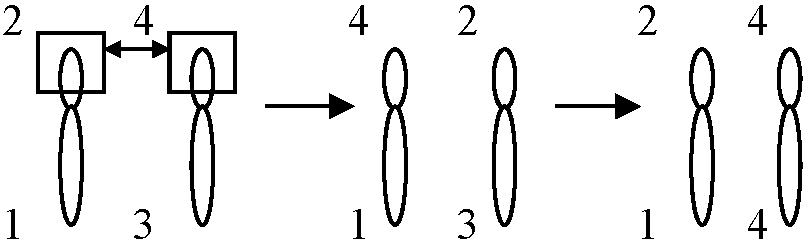 致死遺伝子を防ぐ
致死遺伝子を防ぐ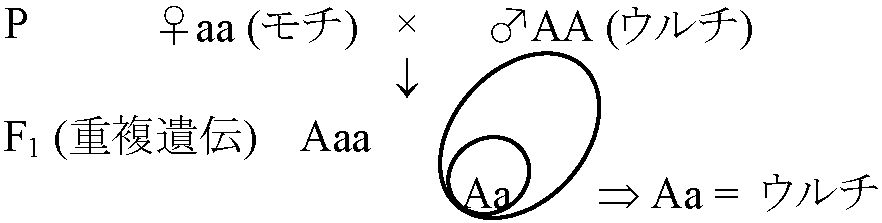

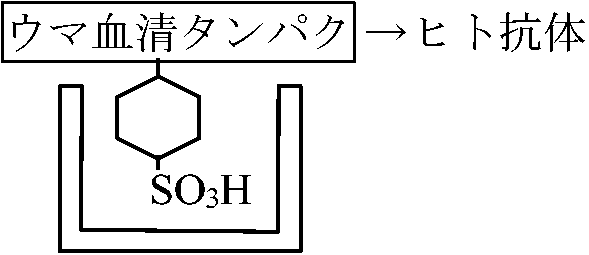 ウマ血清蛋白に対し抗体を作るのではなく、人工的に作ったC6H11-SO3H等に対しても抗体を作る
ウマ血清蛋白に対し抗体を作るのではなく、人工的に作ったC6H11-SO3H等に対しても抗体を作る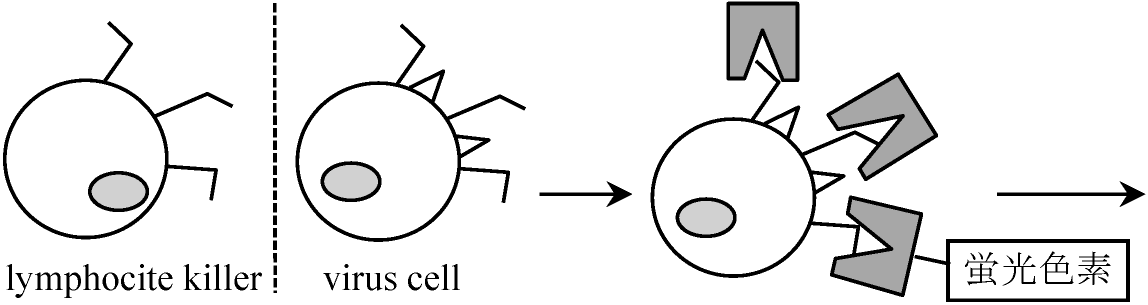

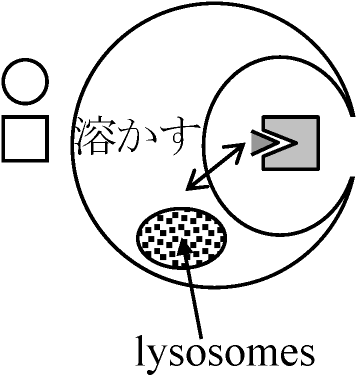
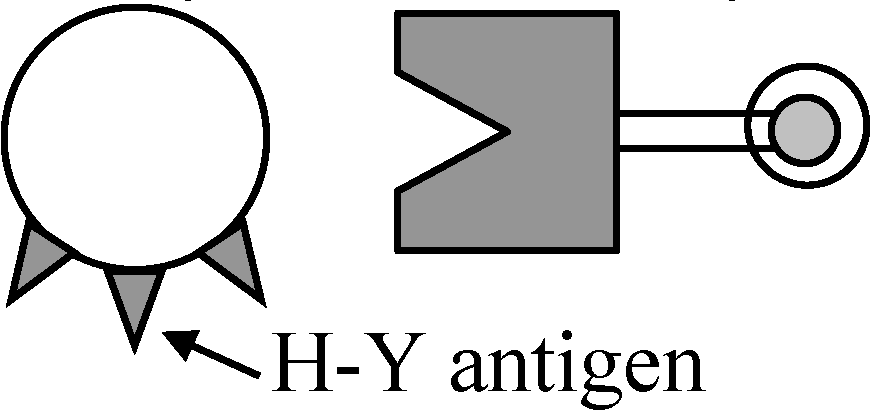
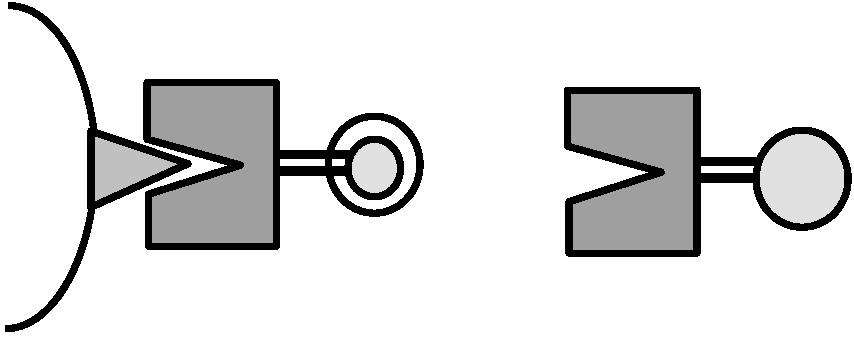
 不溶 ∴ 沈着 ⇒
不溶 ∴ 沈着 ⇒