(2023年9月26日更新) [ 日本語 | English ]
Def. シダ植物: 無種子維管束植物の総称羊歯類 (ferns) と蘚苔類 (mosses)
系統的特徴: 鞭毛 / クロロフィル / 体制 シダ係数 (pteridophyte coefficient)気候係数の一種: (仮定)羊歯植物は高温多湿なところほど種数が多い= (シダ種数 × 25)/(種子植物種数) 1/25を世界標準 (10% = コスモポリタン ↔ 90% = 限定地域 endemic) 北海道 = 1.6, 本州 = 2.1, 九州 = 3.1, 屋久島 = 5.1 羊歯系統分類体系 (PPG)小葉植物 Lycophytaヒカゲノカズラ目 Lycopodiales イワヒバ目 Selaginellales ミズニラ目 Isoetales 大葉植物 Euphyllophyta トクサ目 Equisetales: 分子系統解析 (Grewe et al. 2013, Knie et al. 2015) ハナヤスリ目 Ophioglossales リュウビンタイ目 Marattiales ウラボシ目 Polypodiales |
ヘゴ目 Cytheales サンショウモ目 Salviniales フサシダ目 Schizaeales コケシノブ目 Hymenophyllales ウラジロ目 Gleicheniales ゼンマイ目 Osmundales Note: ナヨシダ, カラフトミヤマシダ - 勇払 生態
分布: 熱帯分布属 = シダ植物 > 17% ↔ 種子植物 ≈ 2% |
採集と観察⇒ 採集: 種子植物と同じ
できるだけ胞子嚢のついた葉(成熟した大きな葉)を含める
小羽片を切り取る → 封筒、薬包紙などに包む → ⇒ 標本作製: 種子植物標本と同じ 前葉体培養(腊葉標本からではなく生葉から)できるだけ新鮮で汚れていない胞子を用意
胞子採取時期は、胞子が胞子嚢から飛び出す直前がよい
(目安: 脳子嚢が黒く色づき光沢持つ段階)
土壌: 胞子から胞子体を育てるのに適 (↔ 前葉体の詳細観察には不向き
素焼きの植木鉢に(畑の)土を入れオートクレーブ滅菌 →
培養のみ目的 - 鉢は水を入れた容器に入れ、埃が入らないよう蓋をし、窓際などの明るい所に置いておけば十分(ミズゴケ培地も同様)
(必要に応じハイポネックス等添加)
寒天(いわゆる寒天培地): 前葉体培養では最も普通
クノップ液、マイヤー液 観察前葉体観察: 顕微鏡 + 解剖顕微鏡(前葉体観察)
スライドグラスに数滴の水を落とし、仮根に絡む土・寒天を除去 前葉体を傷つけないよう注意
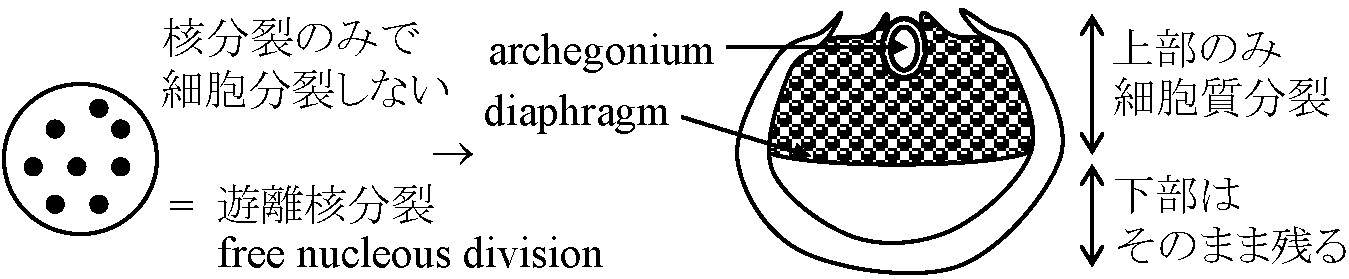
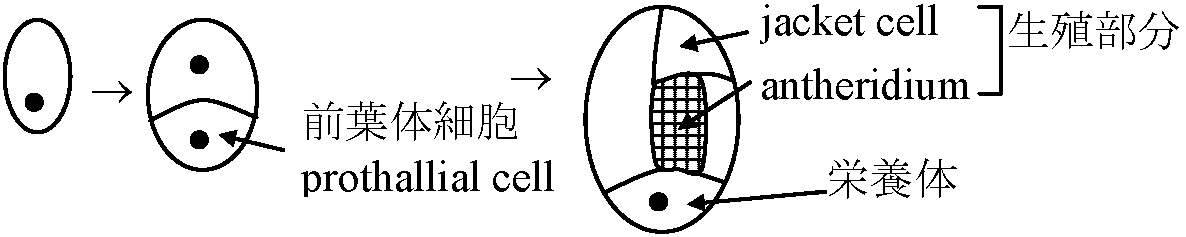
水で封入しカバーグラスをかけ顕微鏡で観察 水で封入すると気泡が入る場合 - 中性洗剤を加えた水を使う 外部形態 external morphology前葉体 ☛ prothallium (配偶体 gametophyte)翼 wing: 前葉体に広がる薄く平たい側部クッション(褥) cushion: 前葉体腹面中央のやや盛り上がった部分 仮根 rhizoid: 前葉体腹面に生える細い糸状構造 (維管束はない) 造精器 antheridium (♂) + 造卵器 archegoium (♀)
+ 付属物 appendage Ex. 鱗片、毛 胞子体一般に観察されるシダの主要形態 ⇒配偶体から発生した2倍体世代 = 根 +茎+ 葉 (胞子嚢群で胞子形成)
鱗片葉: 鱗状に重なる厚い葉
頂羽片 terminal pinna
下向第1小羽片 downward first pinnule ↔
裂片 pinna (pl. pinnae): 羽片ほど独立しないがまとまりある葉片部分 終裂片 terminal pinna: 裂片中で最小 |
Def. 胞子嚢群(嚢堆) sorus: 胞子嚢 sporangium が数個以上集まったもの 縁生 marginal: 葉縁に付く Ex. ゼンマイ、フモトシダ 背生(表面性) laminar: 葉面に付く
頂生: 葉脈先端に付く Ex. シノブ 連続子嚢群 continuous sorus: 胞子嚢群同士が連続し境界不明瞭
帯状、線状、面状
葉裏全体に付く Ex. ベニシダ
機能: 胞子嚢群保護
包膜の付き方
包膜上位: 胞子嚢群を上から覆う様につく
+ 細胞形、色、腺毛、毛状突起有無 側糸 paraphysis (pl. -es): 胞子嚢に混じる不稔の毛状部分 (1/3の種が持つ)
機能: 胞子嚢群保護(乾燥抑制・微環境改善) 内部形態 internal morphology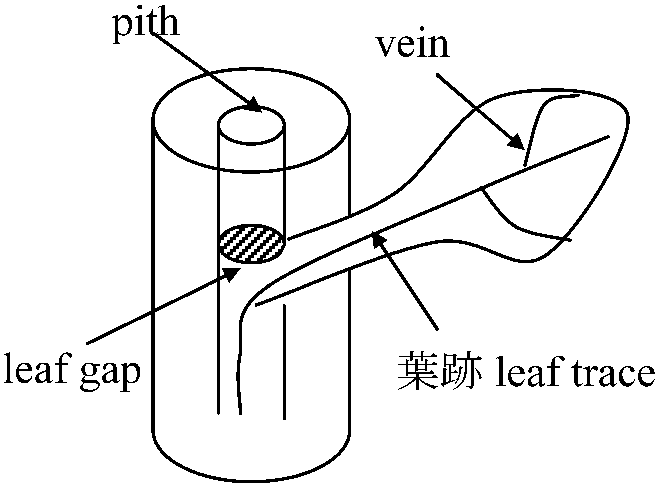
胞子体茎 stem外: 外皮 cortex
柔組織 parenchyma
木部 xylem 葉 leaf 大葉 macrophyllous type = 葉身leaf blade (lamina) + 葉柄leaf stalk (petiole), 葉隙leaf gap形成 高等になるとleaf gap伸びる 小葉 microphyllous leaf: leaf gap無、leaf stalk無、leaf vein 1脈染色体観察固定液: カルノア液、ファーマー液、ニューカマー液
ファーマー液(大抵これ): 99% EtOH + 氷酢酸 |
|
= Gr: Pteridophyte, ENG: fern or vascular cryptogam 単一系統とは考えられず、最低4系統が羊歯植物中に認められる。蘚類苔類レベル以上に大きな特徴で分類されていると言え、羊歯植物と種子植物との差と同程度の分類大系ともいえる
環帯の有無に着目 繁殖器官 (reproductive organ)胞子: 2形態 = 両面体(二面体): 多くは楕円形 + 四面体胞子型 spory a) 同型胞子型 homosporous type (homospory) Psilotum, Lycopodium (トクサは軽い性分化ある) b) 異型胞子型 heterosporous type (heterospory)クラマゴケ、ミズニラ、リンボク*、フウインボク*、デンジソウ、サンショウモ |
配偶体 gametophytes= 前葉体(n) = 退化・小型化 → 異型胞子化に伴う変化a) 胞子内発芽 endosporic germination → 独立 b) 大胞子嚢中で受精: 胞子嚢から胞子が飛び出すか、胞子嚢から出た後に発芽 → 水への適応 sporophyte上生活 ⇒ sporophyte = 大型、優占 胞子発芽 → 前葉体形成
遠心発芽: 仮根と前葉体の伸長方向が反対 生活環 (life cycle)前葉体 (prothallium, = gametophyte): 維管束なく単層構造主体胞子体 (sporophyte): コケに比べ巨大で維管束(vascular bundle)有し生活史の中心となる ⇒ 胞子体をさらに発達させたのが種子植物ということもできる 始源細胞2分
|
|
= or 裸茎植物(マツバラン)門 (Division Psilophyta) 叉状分枝(2叉分枝) dichotomy 原生中心柱 protostele 頂生胞子嚢 terminal sporangium 真性胞子嚢(真嚢) eusporangiate: 何重もの層からなる厚い膜をもち多数の胞子をつける ほぼ茎のみ = 葉は持たず、根ほとんどないか十分に発達していないが菌根形成する(基部に菌類が寄生し菌糸を伸ばしそれが栄養分を吸収する)
維管束植物中もっとも簡単な構造であり、維管束植物の起源と考えられる(Rhynia) 古生マツバラン綱 (Class Psilophytopsida)*化石種 |
マツバラン綱 (Class Psilotopsida)古生マツバランとの系統関係は否定的付属物: 地上茎に螺旋状につく
Psilotaceaeでは刺状 マツバラン (Order Psilotales)退化した小葉 = 小葉類と考えるマツバラン Psilotaceae: 無葉類(プシロフィトン) → マツバラン マツバラン Psilotum L. イヌナンカクラン Tmesipteridaceae 日本: 自生しない Tmesipteris Berhn.: 体制上の枝(付属物)が葉に見える |
|
= 小葉植物植物門 (Division Lepidophyta or Lycopodophyta) 根、茎、葉分化 背腹中心柱 plectostele Mesophyllous leaf → leaf gapなし 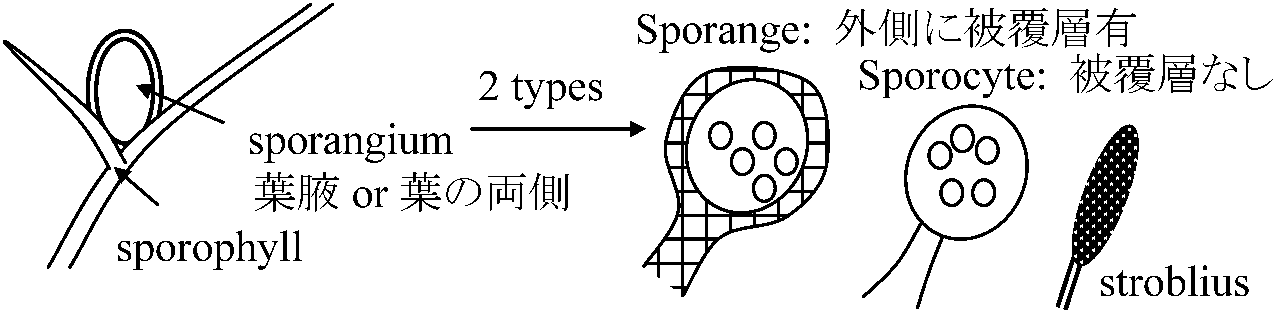 円錐体 strobilus: sporangiumが集まって胞子嚢群となったもの ex. スギナの土筆 同型胞子 homospory: 形成される胞子全て大型同大。一般にgametophyteは雌雄同株(例外 スギナ) 接合子(zygote)発達様式: Archegonium内側に向かい発達 → 種子植物胚は多核体となった後に細胞壁形成があるが、ヒカゲノカズラでは胚は多核体とはならず、直ぐに細胞壁を作り発達する。しばらくは親のarchegonium内で栄養をもらい育ち、やがて外へ排出される 小葉植物綱 (Class Lepidopsida)Glossopsida設ける場合は無舌綱 (Aglossopsida)古生ヒカゲノカズラ* (Order Protolepidodendrales)デボン紀ヒカゲノカズラ (Order Lycopodiales)石炭紀-現世ヒカゲノカズラ Lycopodium L. 有舌綱 (Class Glossopsida)設けずLepidopsidaに統一する見解リンボク* (Order Lepidodendrales)デボン紀-二畳紀種子鱗木 (Order Lepidocarpales)綱設けずLepidocarpaceaeをLepidodendales, MaidesmiaceaeをSelaginellalesとする見解)Lepidocarpaceae*: Ledpidocarpon Scott Maidesmiaceae*: Miadesmia Betr. ミズニラ (Order Isoetales)白亜紀-現世 (世界60-80, 日本2, 道1)Isoetes asiatica ヒメミズニラ) |
葉、根は中央で生じ外側へ押し出される 髄のある茎。葉の基部が残る 茎2次成長する → cambiumがわずかにみられる 春できる葉は栄養細胞のみだが秋になると葉基部に生殖器官できる イワヒバ (Order Selaginellales)石炭紀-現世小葉 (microphyllous leaf) 
小舌ligule: 機能不明。ヒカゲノカズラにはない 胞子母細胞 mother cell (parent cell)
1 → 全て胞子となる = microspore microspore → ♂ macrospore → ♀ 発達様式1. Macrospore (♀)
イワヒバ Sellaginella P. Bequv. |
現世種は全て小葉型 microphyllous type
有節植物綱 (Class Arthropsida)= トクサ綱(楔葉綱) horsetail and sconring rushes根・茎・葉発達。茎葉関節有。葉輪生し、node, internode区別明瞭 胞子嚢茎頂端部に形成。胞子は葉緑体を有し独立栄養 胚発生 embryogamy: (一般) suspensorを有する胚はsuspensorが外側に位置 → 配偶体内側に向かい発達 Order Protoarticulales (Hyeniales)*Order Equisetales (トクサ)石炭紀-現世 (現世Equisetumのみ、染色体数 n = 108)草本的, 二次肥大成長しない  穿孔管状中心柱 perforated solenostele: node部に維管束融合し1つの管 = siphonostele変形(≠ 真正中心柱 eustele)
穿孔管状中心柱 perforated solenostele: node部に維管束融合し1つの管 = siphonostele変形(≠ 真正中心柱 eustele)トクサ (Equisetaceae)トクサ Equisetum L. 極地森林限界越えると観察できない(プルドベイで見た)分類鍵 1. 茎夏緑性、胞子嚢穂つける茎とつけない茎は同形か二形、少なくとも胞子嚢穂つける茎分枝。胞子嚢穂先端円頭 ___ subgen Equisetum (ミズドクサ) 2.茎二形なし、胞子嚢穂つけない茎は規則正しく枝輪生し、つける茎は葉緑体を持たず淡紅-淡褐色 3. 葉鞘0.3-1 mm l(胞子嚢穂つけるもので2 mm)。歯片暗褐色、白膜縁 |
4. 主軸隆条に刺なく、葉鞘断片は僅かに白膜縁。胞子嚢をつける茎は胞子散布後すぐに枯死する ___ スギナ 2. 胞子嚢穂つける茎とつけない茎は同形、共に緑色、疎らに分枝
3. 葉の髄腔は茎直径の3/4以下、表面の隆条は5-10条でやや顕著。葉鞘の歯片の辺縁は僅かに白膜縁 ___ イヌスギナ 2. 茎屈曲せず直立し、髄腔があって中空、主軸 ≥ 径1 mm
3. 茎は節で2-3本の枝を不規則に出す ___ イヌドクサ
4. 葉鞘に黒線あり、歯片早落性。茎太さは2.5 mm以上 ___ トクサ 2. 茎不規則波状に屈曲し、髄腔なくても中実、径 ≤ 0.6 mm ___ ヒメドクサ E. hyemale L. (トクサ)胞子体は胞子出すと枯れ、別の所から栄養体出る
茎に硬い珪酸質の筋 - 物を磨くのに用いた 🧑🍳胞子茎(ツクシ): 節間短く太いのが良い - 頭部と袴(ハカマ)はとる (下処理) 茹でて水に晒し灰汁抜き →
天ぷら、汁の実、スギナご飯、佃煮、お浸し、三杯酢、和え物、卵とじ |
|
分類形質 Ex. 胞子嚢群、包膜、葉、葉柄、胞子嚢、根葉茎、胞子嚢発達様式 葉: 形・色・質
互生大葉型、葉中心にleaf gap 真嚢シダ類 薄嚢シダ類 図. 胞子嚢の付く位置 葉下面か変形付属体につく 胞子嚢 (☛ 図) 根葉茎: 発達 胞子嚢発達様式による区分: 系統群を反映しているものではない a) 真嚢シダ類 (Eusporangiate) = ハナヤスリ目 + リュウビンタイ目
第三紀-現世
1) ゼンマイ科, ウラジロ科 環帯(口環) annulus (pl. annuli): 胞子嚢につく1列の細胞からなる組織。乾燥で伸び温度上がると縮み、その動きで胞子放出。種レベル分類鍵 口細胞 lip cell: ある種シダ類の環帯を中断する薄膜細胞 b) 薄嚢シダ類 (Leptosporangiate): シダ目 + デンジソウ目 + サンショウモ目普通に羊歯というと、この仲間を指すことが多い = 羊歯中種数最多
種子植物全体であればeusporangiumの方が種数が多い
発生初期: 原生中心柱 protostele
胞子穂、実葉 fertile spike (= sporophyll) Sori (pl. sorus): 葉裏側にできる 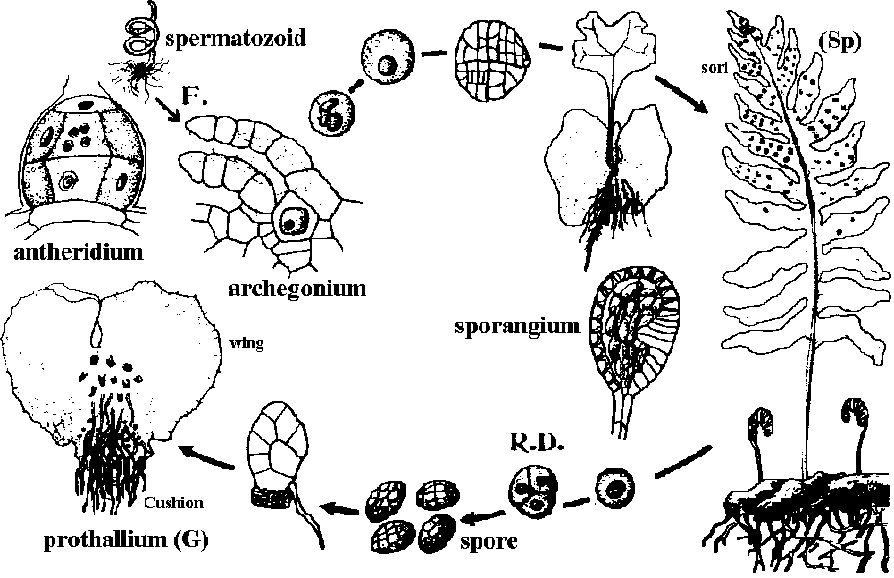 Fig. 羊歯の生活環, Polypodium vulgare (Engler's Syllabas 1954)
前葉体: 葉緑素持ち、寒天培地上で無機培養が比較的容易 成長遅く独立生活に数年必要 - 独立後普通2-3年fertile spikeない 真正シダ亜綱 (Filiciedae)真嚢シダ (Eusporangiales)第三紀-現世Ophioglossaceae (ハナヤスリ): 世界 4 gen. 70 spp Botrychium Sw. (ハナワラビ) Ophioglossum L. (ハナヤスリ) 世界30種 (日本8種 + 1雑種) 染色体基本数: x = 120 検索鍵 (田川 1959) 1. 葉に栄養葉なく、1本の茎に胞子葉のみを5-7本叢生する ___ O. kawamurae Tagawa (サクラジマハナヤスリ) 1. 葉は栄養葉と胞子葉に分岐し、1本の茎に1-3本生じる
2.胞子外皮に粗い網目模様があり、輪郭では瘤状の突起に見える ___ O. vulgatum L. (ヒロハハナヤスリ)
3. 栄養葉は三角状卵形・円形・楕円形ときに広披針形、基部は急に狭くなり、葉柄と明瞭に区別できる ___ O. petiolatum Hook. (コヒロハハナヤスリ)
4. 栄養葉は線形または狭いスプーン形、多少とも先太り ___ O. thermale Kom. (ハマハナヤスリ) 有珠 (原 1978): ヒロハハナヤスリ、コハナヤスリ O. austroasiaticum M. Nishida (タイワンハナヤスリ)O. namegatae M. Nishida et Kurita (トネハナヤスリ) O. parvifolium Grev. et Hook. (イオウジマハナヤスリ) O. parvum M. Nishida et Kurita (チャボハナヤスリ) Ophioderma Pr. (コブラン)本属をOphioglossumから分けるときO. pendulum L. (コブラン) Marattiaceae (リュウビンタイ) 前薄嚢シダ (Protolepidosporangiales)(+ Osumudales ゼンマイ ⊂ 薄嚢シダ目 - PPG廃止) ⇒ 独立目ゼンマイ (Osumudales) = Osmundaceae = OsmundaOsmunda L. (ゼンマイ)O. claytoniana L. (オニゼンマイ) O. japonica Thunb. (ゼンマイ) 🧑🍳5-6月 丸く巻き込み綿毛に覆われた若芽・茎 (乾燥保存は味高める) (下処理) 要灰汁抜 → 和え物、煮物、油炒め、汁の実 O. lancea Thunb. (ヤシャゼンマイ) O. cinnamomea L. (ヤマドリゼンマイ) = ヤチゼンマイ (道で呼ぶ人多) 🧑🍳4-6月 赤褐色綿毛残る渦巻状栄養葉 ☛ 山菜 (下処理) 灰汁抜き必要(木灰か重曹に塩加える) →
汁の実、煮物、芥子醤油、油炒め、和え物、炒め煮 薄嚢シダ Leptosporangiales (= Polypodiaceae, s.l.)石炭紀-現世Pteridaceae (イノモトソウ) 葉縁にsori。Coenosorus形成(肥厚、内捲)。根茎 = 環状中心柱 Pteridium Gled. ex Scop. (ワラビ) P. aquilinum (L.) Kuhn (ワラビ) 🧑🍳開葉前 地上部(灰汁強い) (下処理) 要灰汁抜 →
叩き、汁の実、お浸し、二・三杯酢、油炒め、煮付、蕨飯、和え物、おでん Pteris L. (P. vittata L. モエジマシダ, アジア-米国) Dennstaedtiaceae (コバノイシカグマ), イノモトソウ科と分ける場合) Parkeriaceae or Adiantaceae (ホウライシダ) Adiantum L. (クジャクシダ) A. capillus-veneris L. (ホウライシダ), 葉身3角状長楕円形円頭, 2回羽状 |
Dennstaedtia wilfordii (Moore) Christ (オウレンシダ), syn. Sitobolium wilfordii (T. Moore) L. A. Triana et Sundue): 夏緑生。根茎細く這い軟毛あり。葉ほぼ無毛、やや2型 Hymenophyllopsidaceae (コケシノブ) ≈ 700 spp.(日本29種) 高温多湿な生息地 = 熱帯多 + 着生植物多 葉面小型化、細胞1層 1861 van den Bosch (1812-1870) Gonocormus minutus (Bl.) v. d, B. ウチワゴケ ⇒ 葉多形 ⇒ 4種に区分
G. diffusus (Bl.) v. d. B. ウチワゴケに似るが葉円形・掌状
形態的変異の範囲内であり4種に分けることはできない  図1. シノブ科及び近縁群のrbcL, accD葉緑体遺伝子領域を用いたML系統樹を基にした形質進化系統樹(ACCTRAN) (堤・加藤 2012) キジノオシダ (Plagiogyriaceae) Plagiogyria (Kunze) Mettenius (キジノオシダ), 1属 ヘゴ (Cyatheaceae): Aspidiaceaeに入れる, s.l.) ヒメシダ (Thelypteridaceae) Thelypteris Schmidel ヒメシダ, march fern T. palustris ヒメシダ T. nipponica ニッコウシダ T. phegopteris (L.) Slosson ex Rydb. ミヤマワラビ (syn. Phegopteris polypodioides Fee): 根茎長く匍匐し密に鱗片で覆われた先端部を除き殆ど裸出。葉疎ら、殆ど透明な膜質、全縁 オシダ (Dryopteridaceae, s.s., Aspidiaceae, s.l.) オシダ Dryopteris: 前葉体雌雄同株。普通♂先熟で自家受精しない。embryogamyは外向きに発達 イノデ Polystichum Roth カナワラビ (Polystichopsis (J. Sm.) Holtt.): リョウメンシダ (P. standishii (Moore) Tagawa = syn. Arachniodes standishii (Moore) Ohwi) クサソテツ (Matteuccia Tadaro) ヘラシダ (Diplazium Swartz): キヨタキシダ (D. squamigerum (Mett.) Matsum.), ミヤマシダ似 → 根茎短いかやや株状。葉柄鱗片より狭披針型。小羽片切込み浅く全体に裂片広い Oleandraceae (ツルシダ), separated from Dryopteridaceae, or merged into Davalliaceae) Blechnoideae (シシガシラ) Blechnum L. (ヒリュウシダ) 1. 葉柄及び中軸下部の裏面は暗紫褐色、蜜に細点有りざらつく ___ B. castaneum Makino (ミヤマシシガシラ) 1. 葉柄及び中軸は淡緑色または淡褐色、細点著しくない ___ B. niponicum (Kunze) Makino (シシガシラ)、B. amabile Makino (オサシダ) Asplenium L. (チャセンシダ) Hymenasplenium Hayata (ホウビシダ/ヒメチャセンシダ) Order Gleicheniales (ウラジロ)南極大陸以外の全大陸に分布Gleicheniaceae (ウラジロ), Aspidiaceae (s.l.)に合一 Diplopterygium (Diels) Nakai (ウラジロ) Matoniaceae (マトニア) Dipteridaceae (ヤブレガサウラボシ) Woodsiaceae (イワデンダ), 本科を設ける時にはAthyriumも本科に含まれる Athyriaceae (メシダ) (Aspidiaceae, s.l.に含む) Athyrium Roth (メシダ) Polypodiales (ウラボシ)Polypodiaceae (ウラボシ)日本: 12属50種 主に着生植物 網状中心柱 - 表面には鱗片並ぶ 胞子嚢群: 包膜なし ノキシノブ Lepisorus (J. Sm.) Ching Vittariaceae (シシラン) Vittaria Sm. (シシラン) ☛ largely moved to Haplopteris V. flexuosa Fée (シシラン)✂, south to Kanto V. fudzinoi Makino (ナカミシシラン)✂, south to Chubu V. zosterifolia Willd. (アマモシシラン)✂, south to southern Kyushu Haplopteris C. Presl (シシラン, when established) H. anguste-elongata (Hayata) E. H. Crane (ヒメシシラン), syn. V. anguste-elongata Hayata H. ensiformis (Sw.) E. H. Crane (ムニンシシラン), syn. V. ensiformis Sw. H. flexuosa (Fée) E. H. Crane (シシラン) H. fudzinoi (Makino) E. H. Crane (ナカミシシラン) H. mediosora (Hayata) X. C. Zhang (イトシシラン), syn. V. mediosora Hayata H. taeniophylla (Copel.) E. H. Crane (ヒロハシシラン), syn. V. taeniophylla Copel. H. yakushimensis C. W. Chen et Ebihara (オオバシシラン) H. zosterifolia (Willd.) E. H. Crane (アマモシシラン) H. flexuosa (Fée) E. H. Crane × H. fudzinoi (Makino) E. H. Crane (セトシシラン), syn. V. flexuosa Fée × V. fudzinoi Makino 水生シダ (Hydropteridales)白亜紀?、第三紀-現世⇒ Marsileales/Salviniales分割見解
共通点: Sporocarp = sorus: indusiumに包まれる Marsileales (デンジソウ), 設ける場合Marsileaceae (デンジソウ)Marsilea L. (デンジソウ): 田字草 → 四葉 quadrifolia Sporocarp発生: 羽片起源 (Purid & Garg 1954) ↔ 葉起源 (Gupta 1962) Solenostele: Leaf gap = macrophyllous / Sporophyllが折り畳まれsporocarpになる Heterosporic (異型胞子性): 基部 = macrosporangium, 先端部 = macrosporangiumM. quadrifolia (デンジソウ): 水田や水位変動少ない池等。落葉。細根茎泥中這う。4枚の小葉からなる葉を水面に浮かべる。道-九州) Salviniales (サンショウモ), 設ける場合Salviniaceae (サンショウモ)Azollaceae (アカウキクサ) Heterosporic。胞子形成および発芽形態は前2種と同じ |