(2024年7月22日更新) [ 日本語 | English ]
HOME > 講義・実習・演習一覧 / 研究概要 > 小辞典 > 種
種数 (species richness)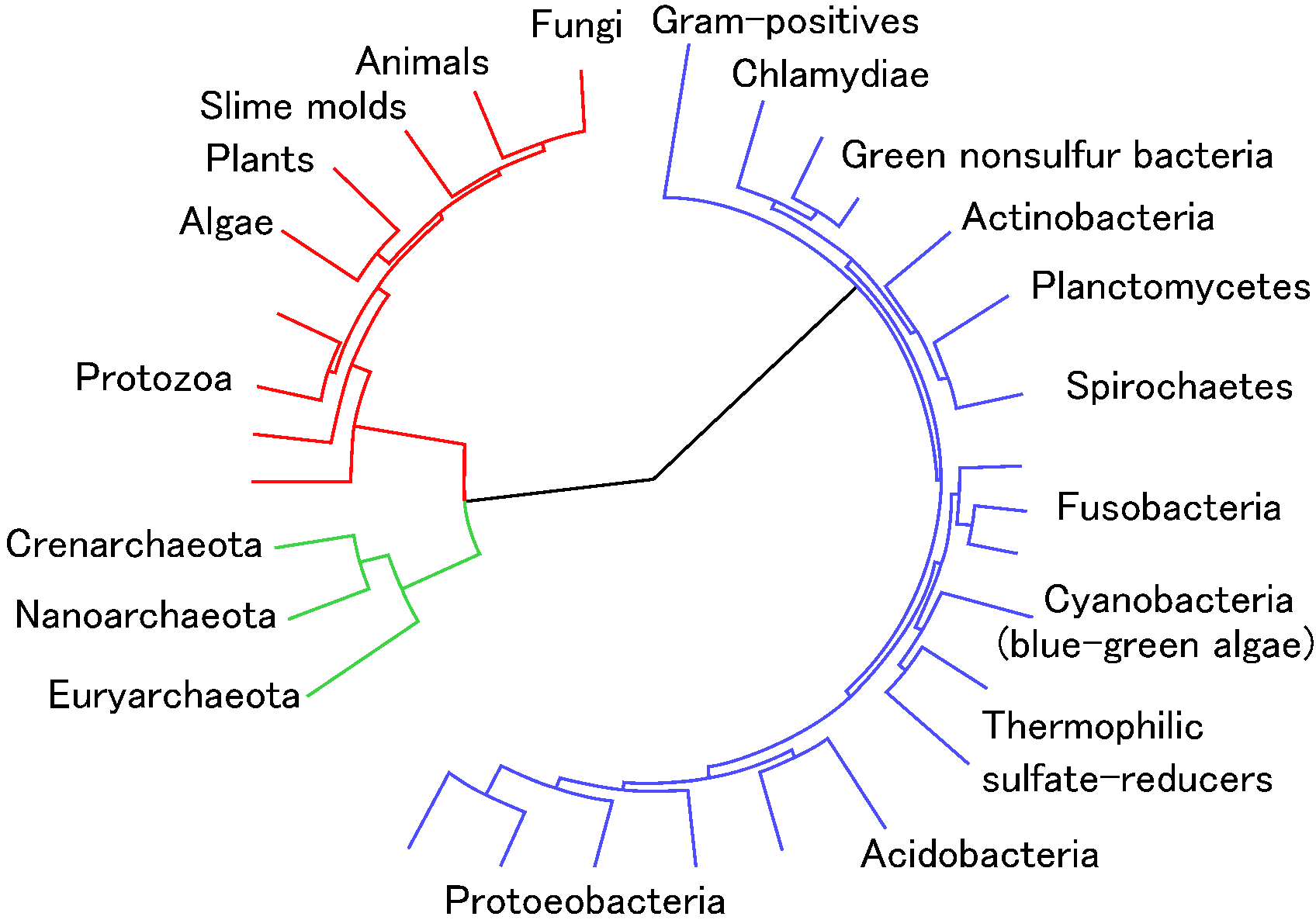
共通祖先に起源するとしたときの遺伝子解析をもとにした現世生物分類群の系統樹 (phylogenetic tree) Euryarchaeota (ユリアーキオータ, 古細菌), Nanoarchaeota (ナノアーケオタ, ナノ古細菌), Crenarchaeota (クレンアーキオータ, 古細菌の一部), Protozoa (原生動物), Algae (藻類), Plants (植物), Slime molds (粘菌), Animals (動物), Fungi (菌類), Gram-positives (グラム陽性菌), Chlamydiae (クラミジア), Green nonsulfur bacteria (緑色非硫黄細菌), Actinobacteria (放線菌), Planctomycetes (プランクトミケス, グラム陰性菌の一部), Spirochaetes (スピロヘータ, グラム陰性菌の一部), Fusobacteria (フソバクテリウム), Cyanobaceria (藍藻), Thermophilic sulfate-reducers (硫酸還元菌), Acidobacteria (アシドバクテリア, グラム陰性菌の一部), Protoeobacetria (プロテオバクテリア) |
[エングラー体系 (Engler's syllabus)]
生物種 種数 生物種 種数 生物種 種数
ウイルス 1,000 藻類 26,900 原生動物 30,800
細菌 3,000 緑藻類 7,000 動物 1,033,614
シアノバ 1,700 褐藻類 1,500 海綿動物 5,000
クテリア
菌類 46,983 紅藻類 4,600 腔腸動物 9,000
子嚢菌類 28,650 その他 14,400 扁形動物 12,200
担子菌類 16,000 植物 248,428 線形勤物 12,000
真正粘菌 500 コケ類 16,600 環形動物 12,000
細胞性粘菌 13 シダ類 10,000 軟体動物 50,000
その他 1,820 裸子植物 529 棘皮勤物 6,100
被子植物 220,000 節足動物 874,161
その他 1,299 昆虫類 751,000
その他 123,161
脊推動物 43,853
その他 9,300
合計 1,392,425
系統進化学の手法形態学
比較解剖 Ex. Paeonia: Ranunculaceae → Paeoniaceae 細胞学・遺伝学
染色体数・核型分析・ゲノム分析
二次代謝産物 |
種とは What is species?= 分類学(植物分類学, plant taxonomy)のメインテーマ生物界: 構造単位が順次高次システムにまとまる → 種species単位存在 = 基本概念: 階rankと類型category 生命の存在様式 = 名前a) 確かな名前 名前:対象物 = 1:1 → 曖昧な意味なく厳密な対応関係b) 名前の整理 c) 名前の種類 学名 scientific name, 和名(一般和名) Japanese common name 記載的研究 monograph: あるグループにどのような植物があるのかを研究 植物相的研究 flora: ある地域にどのような植物が生息しているのかを研究(動物 = fauna) 種の存在様式 = 個体各レベルごとに多くの相違点と適用限界があるが、少なくとも3点で基本的共通性存在1. 各々の構成レベルにおける内的な統一性存在 2. 各々のレベルで再生産する能力を備える 3. それ自身が時間の経過と共に変化する 1. 分類学的種 taxonomic(al) species= 形態種 morphospecies) - morphological species concept[仮定] 外界環境が変化しない → 標徴(記載)に与えた特徴を備えた全個体は同一種に属す(Plate 1914) → 類型的種概念 typological species concept (international nomenclature) リンネ種: Linneon = 形態種 morphospecies → 基準標本(タイプ標本)
実用上の必要性 → 使うのに便利で実用的convenient and practicalな分類でなくてはならない 種は全分類群上特別な位置がある? / 種以外の分類群は抽象的概念? |
2. 生物学的種 biological species= 自然種) → biological species concept (高等動物分類での導出概念)生物学的種 = "自然状態"では交配しない(Mayr 1969) 「個体の遺伝子が世代を通じ伝わり、一個体中に合体することが自然条件下で可能なら同種」 基本的に構造・機能同一であり、有性生殖により生殖能力のある子ができる 生物学的種に対する批判 生殖隔離は実際は存在しない → BSでは種認識できない(現在否定) → 生物学的種概念は、その間に生殖的隔離の手段を問わずバリヤーの存在を区分として認めようというもの (Imbrie 1957) 3. 遷移種 successional species
4. 生態種 ecospecies= 生態型をまとめて一つの種とするエコタイプ (生態型, ecotype)種レベルで複数タイプが環境に対応した関係が認められる → 複数エコタイプEx. Hieracium umbellatum (Turesson 1922, 1925)
4形態(砂丘・非移動砂丘・崖・森林) - 栽培で形態変化しない・交雑可能 コンビビウム convivium: 自然交雑するが形態地理的区別可能な個体群 コンミスクウム commiscuum: 実験的には交雑し稔性ある子孫残せる個体群 コンパリウム comparium: 雑種を生じる限りの個体群 エコクライン (生態勾配, ecoclinee)環境勾配に沿った生物の(遺伝的)形質の変化形質 (character)≈ 遺伝学でいう形質 character遺伝子の具現化したもの → 形態が遺伝子差を表す Biological species判断 → 形態種を見ることが有効な手段 元々は分類指標となる形質的要素を指し、その意味で特徴(標徴)とも呼ぶ。メンデル以来の遺伝的形質を指し、その内の単位的形質、即ち単位形質を捉えて遺伝現象を分析することが遺伝学の常套手段の一つとなった |
種分化を論じる上での概念的用語Formenkreis型圏 (Kleinschmidt 1900): 最初動物学で提唱された種を論じる上での基本単位合弁花 > 子房下位 > 雄蕊5個 … ? → これ以上分けられない種の共通単位 Rassenkreis oder Artenkreis (Rensch 1929) 最初動物で提唱同一祖先と思われるものの集団。species complex (super-species)の起源的用語 Polytypic species (Huxley JS 1940) 動物 ↔ monotypic speciesSuper-speciesに通じる。形態は異なるが共通的種とみなせる集団 形質転換 (character displacement)似たような種がいない場合には大きな変異 ⇔ 類似種が近くにいるsympatric場合交雑をさけるため形質特異化
交尾前の隔離機構の一つ – 強化 reinforcement 交配前隔離異所的隔離 allopatric isolation: 近縁種が地域的に離れて生息 ⇔同所的隔離 sympatric isolation 交配後隔離交配後隔離機構での変異 variation: 染色体変異 Ex. マイマイガの性品種⇒ これらはいずれも種分化の一要因と考えられている 変異 (varation)1942 Mayer E
Geographycal race - 動物 (≈ 生態型): 鳥の変異を調べ、島での鳥のraceを考えた → Huxley 遺伝子レベルでの環境が植物に与える影響を考慮すると、環境条件にり起こりうる交雑がある Ex: Pinus densiflora (アカマツ, 西日本): 原植生 = 照葉樹林
↑ introgression Pinus × densi-thunbergii (アイグロマツ) 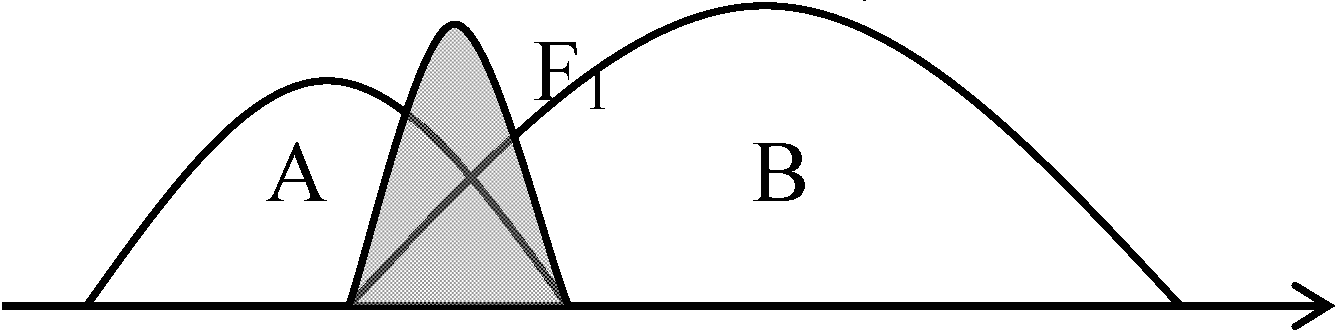 F1 = 不稔 → A, Bどちらかとback-cross可能なことがintrogressionの条件 ↓ 仮にAとback-cross 雑種子孫がAに浸透していくことで雑種起源の新品種形成がなされる 1939 Clausen U: 細胞遺伝学 (Turessonの弟子、Hagerupは兄弟弟子)
Turesson学派は一貫し生態型研究。Stanford大カーネギー研究所Prof Babcockのもとカーネギー財団の「自然と生物の変化パターン」という提出テーマでStebbins, Ravin, Lewis H, 下斗米、田中隆荘ら研究 レース (連携群. race)地理的レース geographical raceslocal racesとの区分は便宜的地理的レベルでの多系 ≈ cline (分類学上geographical raceはsubsepcies程度に相当) 不連続地理的レース disjunctive geographical raceEx. 高山帯等の隔離された個体群 – cp. ecological geographical race 生態的(地理的)レース ecological (geographical) race同一空間にパッチ状あるいは桝目状に形成された個体群の発達 山岳地帯にいおては標高レースaltitudinal raceが発達する (Huxley 1938) クライン (cline)個体群のある形質が地理的環境の変化に応じて連続的に変化する現象
種間クライン: 種内の地理的変異 南北cline: body size factor = 北 ← 寒 → 体重当り表面積 → 小 Ex. 脂肪貯蔵のため体大きい 東西cline: appendage factor = 西部乾燥 → 表面積/単位体重 → 小 Ex. 短足 Ex. Juniperus verginiana (Flake, Rudloff & Turner 1969)テルペンtelpeneをガスクロ分離定量 – 個体群地理的変化に沿う変化 Ex. Trifolium repens: シアン水素配糖体の遺伝子集団分布メンデル遺伝多型 Ex. Mimulus (Becks 1962)地理的レース(クライン)は光周性、光合成特性等にも存在 Ex.Ray & Alexander (1966): オナモミ = 短日性で暗期間に対し感受性
一般に限界暗期は8.3-8.5 hr → Los Angelesでは11-12 hr(1959)
花芽形成が見られたときの暗期をmanifest night-lengthとする 3) natural-day experiment: 適応確認に42.3°N (arboretum)で行なう
暗期に対する感受性sensitivityはHawaiiのもので低い
自然状態natural dayでの花芽形成は、温度の影響を受け、特に8月中旬以降の日長で花芽形成するものは、種子形成までの期間を完結できないものがある |
極地への移入戦略 strategy for migration of polar sites
短日植物 → 長日(あるいは中日)植物への変化 雑種指数 hybrid index
Charater#
この指数は、選択形質数、遺伝子形質との関係、レースの問題等がある レース形成に関与する淘汰要因 selection factors on race formation1922 Turesson: レース区分1. 生育地における優占的な環境要因への適応 Ex. Atriplex litorale: 風の強い生育地と弱い生育地への分化 2. 種間についても置き換えがある
Ex. A. litorale, A. sarcophyllum, A. praecox: 住み分け → 気候による生態型分化(開花期淘汰圧強) BN: wild – BN中でもたまたま農場にあるものがあるものはBC様に特殊化している Ex. Gilia splendens: 受粉問題 ツリアブにより受粉されるレース / ハチドリにより受粉されるレース = pollination race レース形成速度Ex. Mimulus guttatus (Vikey 1978): Lake Bonneville, Utah
湖の窪地での乾地化(= 植物侵入が可能になった)年代が分かる 隔離機構 isolation mechanisms1) 交配前隔離機構 pre-mating isolation同所 的sympatric space / 異所的 allopatric / 側所的 parapatric生態的隔離機構(普通不完全)
Ex. spade food toad: 異なった土質を好む
Ex. ホタルの求愛行動: 擬態で♀を呼び食べてしまう → 発光パターン変えれば交雑発生
番作らない鳥では一瞬で見分けるため形態的にmale羽型が大きく異なる
分布 食相 大きさ 翅鞘
コブN 札幌以北4型 アザミと 中間 変異大
ルイヨウ
コブS 渡島以南 アザミ 大型 もり上がる
ルイヨウ 渡島以南 ルイヨウ 小型 もり上がらない
安定
コブSとルイヨウ: 大きさ、翅鞘異 → 生殖的隔離 = 同所だが食相差異のため交雑しない → 異種 交配後障壁F1発生しない ☛ 種間雑種 interspecific hybridRana pipiens × R. sylvatica: 嚢胚後期に発生停止 - 自然中で生存不可 Ranunculus villarii (marsh) × R. dissectiofolius (dry habitat): F1はどちらの環境でも生存不可
中間の不安定生息地に生存し、すぐ死ぬし生存域も限られ個体数少 生物学的種概念の適応外生物1) 無性生殖(有性生殖だから隔離が可能) Ex. ワムシ2) 種分化途中にある種 = 半種 semi-species 個体群間変異 ≈ 地理的変異 半種 (semi-species)biological speciesとraceの中間的な生殖的隔離が見られる1. 漸進的分岐による 漸進的分岐graducal divergence
early stage (intergradation) = geographical race, local race
↓↑ 中間ステージ = semi-species (Grant 1963) a. 雑種群 b. 浸透性交雑個体群 それまで生態的隔離が起こっていたのが浸透性交雑を行なうようになった(要再考) (Markert & Moller 1959) アイソザイム (イソ酵素, 同位酵素) isozymeDef.古典: 電気泳動で識別できる酵素の変異群単一遺伝子型アイソザイム unigenic isozyme: 同一遺伝子の1次産物であるペプチド鎖の細胞内存在様式の差異による → 様々な生物で見られる
会合型アイソザイム aggregational isozyme: ペプチド鎖の会合度の違い
基質同じくする酵素種の分子多形 Genetic identity index (GII)
Ex. Tragopogon: 種内 = 0.97-1.00 (locality間比較), 種外(種間) = 0.50-0.62
→ この2種は極めて種として近い(現在分化の最中? F2は不稔) アロザイム allozyme1遺伝子座の異なる対立遺伝子によりコードされる酵素のうちの1型 |
☛ 雑種命名規約
種内雑種 intraspecific hybridEx. イノブタ boar–pig hybrid: ブタと比べ肉質良、高耐病性、多産
= イノシシ Sus scrofa × ブタ S. s. domestica* (F2形成可能) 種間雑種 interspecific hybrid異種間での交雑により生じる雑種第一世代(= 異種間雑種)Ex. ラバ mule、レオポン leopon、ライガー liger 雑種弱勢 hybrid weakness種間雑種が両親より低い成育示す - 次世代得られず育種上障害不安定 = 不稔性/不妊性多 ∵ 生殖細胞減数分裂時に染色体の対合や分離が正常に起こりにくい 雑種に倍数化起こると新種形成することがある品種改良 breed improvement生物放射線 ☛ エンレイソウを用いたX線照射実験
遺伝子突然変異 + 染色体突然変異 chromosome mutation
「鬼が出るか蛇が出るか」的アプローチ |
☛ 生命倫理学 動物人工授精 artificial insemination: 採取精液を雌生殖器内に注入し受胎試みる家畜・水産動物等の繁殖・品種改良、ヒト不妊症対策 人工授精: 精子を♀生殖器内部に注入 - ♀体内で授精受胎体外受精(人工受精): 精子卵子取出し受精 - 受精卵を子宮内に戻し妊娠 植物人工授粉 artificial pollination作物改良 ☛ アブラナ, コムギ, トウモロコシ (自然交配もある) 接木 grafting: 栄養雑種(接木雑種)組織培養 tissue culture ☛ 無性生殖 胚培養・葯培養 (n) ↔ 体細胞培養 (2n) 胚培養 embryo culture: 受精後の胚を培養し個体へ育成させる方法
雑種: 胚成長が乳胚発達不全により損なわれること多
1964 Guha &Maheshwari: Datura metel葯培養成功 細胞雑種 cell hybridDef. 複数種細胞を融合させ得た細胞応用: 品種改良、遺伝子治療 |