(露崎担当分 2019年11月14日更新) [ 日本語 | English ]
[ 99 | 00 | 07 | 09 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | PV ]
処分するのはもったいないレポートを掲載しておこう(要は、もったいない)。レポートの内容が正しい、というわけはないので誤解なきよう。名前は、載せても良いのかもしれないけれど、無名ということで。なお、ワープロ提出と手書きのものは、伊藤さんにタイプして頂いた。劣悪なレポート課題を出したため優秀レポートがないか、試験にした年にレポートはないため掲載できない年もある。
明らかな誤字・脱字、個人的に好きではない漢字は、書き直してある。
[英語(一部英語みたいな)レポートはこっち (読めるのだけ)]
エコパン (レポートではないが)最終発表意見以下の意見を今後の発表の場面で活かしてください。 # 似た意見は近くにまとめたつもりですが完全ではありません。 # 表現を一部変えたところがあります。 # 教員の批評を全部フォローできませんでした。分かった分だけ【 】で示しました。 キャンパス班
|
|
環境保全学特論論文の要約Johnson, E.A., Miyanishi, K. (2008) Testing the assumptions of chronosequences in succession. Ecology Letters 11:419-431.多くの生態学の入門的な教科書では、空間を時間に置き換えて遷移の進行を決定する(クロノシークエンス)、古典的な研究を使って遷移を紹介している。この手法には過去に批判があったにもかかわらず、たいていの場合は無批判に、現在でもクロノシークエンスは遷移に関わるトピック(生物多様性、生産性、栄養循環などの時間的な変化)の研究で使用されている。クロノシークエンスを基盤とした研究の問題を一般的に示すため、私たちは4つの古典的な遷移研究における空間から時間への置き換えを、クロノシークエンスではない手法(例えば、永久調査区の長期研究や花粉学、林分の復元)を用いて検証した研究から得られた証拠を総括した。いくつかの例では、検証に同じ場所が使用され、そのうちの一つが同じプロットを使用していた。私たちは実証的な証拠が、古典的な研究で推測されてきたクロノシークエンスを基盤とした結論の説得力を弱めることを示した。(397字/400字) Walker, L.R., Wardle, D.A., Bardgett, R.R., Clarkson, B.D. (2010) The useful of chronosequences in studies of ecological succession and soil development. Journal of Ecology 98:725-736. クロノシークエンスは、複数の時間スケールをまたがる植物群集と土壌発達の時間的な動態を研究する上で、重要な手法である。しかし、多くの場合、不適切に使用され、生態学的なパターンやプロセスについて誤った結論を導く。そこで、私たちはクロノシークエンスを用いるのに適当なのはどのような場合かを評価した。異なる年代の場所が同じ道筋をたどる証拠がある場合には、数十~数百年の時間スケールの植物の遷移を研究する上で適切な手法である。また、数百~数千という時間スケールで生じる土壌発達に関する研究でも信頼して使用できる。種数や植物の被覆、植生構造、土壌有機物の蓄積などの変化は種構成や個体数に比べて予測しやすく、時間的に線形な挙動と関係するため、適している。クロノシークエンスが最も適しているのは、遷移の道筋が収束し、生物多様性が低く、種の入れ替わりが早く、攪乱の頻度や強度が低い群集に関する研究である。(393字/400字) |
相違点の論理的な解決Johnson & Miyanishi (2008)は遷移研究の初期の研究を総括し、「どの遷移年代の場所も異なるのは年代だけであり、非生物的・生物的な構成要素について同じ変遷をたどる」というクロノシークエンス法の仮定は成立しないことが多いことから、クロノシークエンス法を闇雲に使用することに対して警鐘を鳴らしている。一方で、Walker et al. (2010) はクロノシークエンスが妥当な結論に結びつきやすい研究対象に対しては、むしろ積極的に使用してもいいのではないかと議論を展開している。この表面上の相違は、Walker et al. (2010) が指摘しているように、研究時間スケール、空間スケール、生物指標の違いによって生じていると考えられる。例えば、ある地域で攪乱を受けた100 m2の場所の種構成を10年間調べるのと、攪乱を受けた10000 m2の場所の植物量を100年間調べるのでは、遷移の研究でも大きく意味が異なる。短期的・局所的な生物の応答には、近傍の植生や地形の影響なども大きく影響すると考えられるが、長期的・広域的な反応では局所要因の差は小さな問題となる場合がある。また、ある時間スケールにおける種の反応や構成する種構成が異なる場合であっても、種数や生物量といった指標では差が出にくいだろう。また、目的の違いも議論の展開の相違に繋がっているだろう。遷移のメカニズムを特定することが目的ならば、クロノシークエンスには限界がある。多くの攪乱された場所でA種の後にB種が定着していることが確認できた場合であっても、A種が定着したことによってB種が定着できたかどうかは分からない。しかし、A種の後にB種が定着するという現象は、同じ地域の次の攪乱でも見られる可能性は高い。クロノシークエンスは広域的・長期的な大まかな傾向(パターン)を見る上では有効なツールとなり得るが、局所的・短期的な変遷の予測、あるいは遷移のメカニズムの特定に対して使用することはできないと考える。(774字/800字) |
自然環境学総論基礎的な内容よりも、最前線の話題に重点を置くべきだと思います。基礎的な内容であれば、参考書や専門書に記載されています。図書自体は図書館などで借りられるので、自習で十分賄える範囲なのではないでしょうか。一方で最前線の内容は、論文に記載されている事はあっても、本には記載されていない事が多いです。自分で論文を探すのも良いのですが、最前線のものとなると、自分の専門分野以外のものは探しにくいように感じます。基礎的な内容を復習しながら、最前線の話題に触れる事ができれば最上です。ただそうするには、今までの講義を受けている限りでは、かなり難しいという印象を受けました。再沿線の研究については講義で重点的に扱い、基礎的な知識については、受講する学生がすでに学習している前提で良いと思います。 環境保全学特論絶滅が危惧される希少種と、広く分布している近縁種の間で繁殖特性の違いを比較した論文を2つ選んだ。両方の論文では、繁殖特性の違いに注目して、ある種が希少種となりうる原因を探り、その希少種をどのように保全するべきかを考察している。
結果: 花生産と種子生産はD. rotundifoliaの方が大きく、娘生産はD. anglicaの方が大きかった。ただし、2種間で結実率に差はなく、花生産の違いが種子生産に差をもたらしていた。D. anglicaにおいて、花生産後の生存率低下が大きく、娘生産は生存率を増加させた。 結論: 多年生植物には以下の2つの繁殖戦略があり、個体群維持に影響を与えうる。
結果: G. althoferorumと他の2種との間で、繁殖特性に大きな違いが見られた。G. althoferorumでは、繁殖能力のある花粉の生産が少なく、受粉と受精成功が低かった。これにともない、G. althoferorumの果実生産は低く、一つの個体群では果実を全くつけなかった。埋土種子もG. althoferorumの下には全く見られなかった。 結論: G. althoferorumにおいて、両方の個体群で有性生殖による個体群の回復はほとんど見られず、一つの個体群では主に無性生殖によってのみ繁殖していた。人間活動の結果、成体が枯死し加入個体が減少することにより、個体群が縮小しているといえる。このことから、G. althoferorumの保全を行う上で、不慮の破壊から成体を保護することが重要であると考えられる。 |
比較: 私が注目した相違点は、結論に見られる保全方法の違いである。これは、保全対象となっている植物の特性が異なることによって生じる。両種とも、繁殖戦略として、無性生殖によりその土地に固着する戦略をとっているため、成体を保護することは重要である。ただし、D. anglicaにおいては研究により、生育に適した環境が限られていることがわかっている。そのため、D. anglicaを保全する際には、成体を保護するだけではなく生育環境を整えることも大切である。G. althoferorumは、近縁種2種と異なり、潜在的な結実能力がかなり低く(花粉の多くが繁殖能力をもたない)、埋土種子の生産能力が低かった。さらに、人間活動により成体が枯死することにより、無性生殖による加入個体の供給が減ることが、G. althoferorumが希少種となっている原因であった。このことから、G. althoferorumの保全を行う際には、成体の保護がより大切となるといえる。また、この論文ではG. althoferorumの種子生産能力、蓄積された埋土種子の量は調べているが、無性繁殖がどの程度行われているのか、あるいはどのような条件で増加するのかは調べていない。成体の保護のみでよいのか、適した生育環境はあるのか、あるいは他に希少種となる要因はないのか、調べる必要があると考えた。これまでの研究から、競争に弱い(Walck et al. 1999)、散布能力が低い(Fiedler 1987; Hanski and Gilpin 1997)、自殖率が高い繁殖システムをとっている(Karron 1987; Kunin and Schmida 1997; Lavergne et al. 2004) 種が、環境の変化を受けやすく、希少種となりやすいことが明らかとなっている。また、希少種において遺伝的多様性が低いことも、一般的となっている (Cole 2003)。おそらく、D. anglicaとG. althoferorumの2種は無性生殖に依存しているため、遺伝的多様性が低く、環境の変化に適応する能力も低くなっていたと考えられる。一度個体数が減少すると、遺伝的多様性はより損なわれ、その個体群の維持は困難になることから、希少種となる前に個体数減少を防ぐことが重要であると考えた。 引用文献
エコパン [考察のみ]
[考察のみ]道路からの距離が遠いほど、土壌硬度が小さくなると考えたが、結果はそうではなかったので意外だった。恐らく、生息している植物の種によって、土壌硬度が変化しているのではないかと思う。特に、木の下では落ち葉等を微生物が分解して、より土壌が柔らかくなるのではないだろうか。 また今回、照度や土壌硬度から、生息する植物の種に傾向が現れるのではないか、と考えていたが、ある程度、そのような傾向を垣間見ることができた。エゾヘビイチゴでは、照度の比率が高いところほど、全体に占める割合は大きかった。逆に、ナガハグサは、照度の比率が低いところのほうが割合が大きかった。さらに、ナガハグサは照度に関係なく、どこもほとんど同じ高さだったので、少ない光でも繁殖することができると考えられる。オオバコでは、土壌が柔らかく、照度が低いところで割合が大きかった。また、セイヨウタンポポはあまり傾向が見られなかった。これらのことからやはり、群落動態には様々な要因相互に関わっていることを痛感した。 種の多様性に関しては、区画5が最も多様であるといえるが、今回区画5で特別何かの値が変化していることは無かったので、やはり、調査したこと以外の要因が関わっていると考えられる。 今回、やはりサンプルの量が少ないことに加えて、調査する内容も他にいろいろあったのではないかと思うと、色々心残りである。また、種名のみで考察したが、科や属といった枠組みで考えると、また別のことが見えてくるのではないかと思った。また、季節ごとに群落動態も変化すると考えられる。これらを考慮すればもっと面白いデータがとれるのではないかと思った。今回、調査の方法はもちろん、様々な種類の植物の種を見分けることができた。今後、これを生かして、普段からどのような植物が生えているのか注視しながら大学や道を歩きたいと思う。 |
自然環境学総論環境科学の最前線の話を中心にすべきだと考える。本講義はオムニバス形式で各教官4回程度の講義しかできないため、基礎的な内容を話すだけで講義が終わる可能性が高くなる。私は他学院から受講しているので、各教官の専門に特化したこの講義では基礎的な内容でも知らないこともあったのだが、地球温暖化に関する講義は他にも受けてきたため、他の講義とかぶる話も多かった。知っている話ばかりだと、この講義を受ける意味や新しい知識を得る楽しみがなくなるまたは減ってしまうと考える。また私は環境科学が専門ではないので最前線の話はよく知らないし、3分野の最前線を一度に学べる機会はそう無いと思われる。そのため、それぞれの分野の最前線に特化することで他の講義と一線を画し、少ない講義回数でも学生の知識や学習意欲を高めることができると考える。環境保全学特論Uesaka, Tsuyuzaki (2004)北海道駒ヶ岳において、共生する植物に対する落葉樹、常緑樹、裸地パッチの効果を検証した。種多様性はヤナギ、シラタマノキ属のエリアで増加したが、シラタマノキパッチでは増加が遅く共生する植物のシュートの密度も低下した。 シラタマノキパッチの光強度はヤナギパッチより低く、ほとんどの種類の発芽定着を制限したが、これは2つのパッチを構成する種の茎と葉の構造、フェノロジーの違いである。火山灰堆積層の水分と栄養段階、光強度、種の捕捉の改善によってヤナギのパッチは特にヒメノガリヤスにとって定着促進場所として働いた。ミヤマヤナギのような落葉樹パッチは厳しい環境下で遷移の割合を促進させる改善の役割をもち、常緑樹パッチでは植生の定着は遅くなることが示唆された。 Titus JH, Bishop JG. (2014) 部分的な日陰と土壌撹乱処理を施した4つのハビタットタイプ―やせ地、若いLupinus lepidus、老いたLupinus lepidus、の藪―にベイマツの種を植え、生存を観察した。ベイマツの発芽と生存はやせ地と若いルピナスサイトで最も高く、老いたルピナスサイトとアルナスパッチでは低かった。発芽と定着はやせ地と若いルピナスのパッチの日陰と初期土壌撹乱によって高められた。やせ地と若いルピナスのサイトの日陰で上昇した種の生存は、それらのサイトでの土壌水分をめぐる競争の減少に起因するものであり、古いルピナスとアルナスのサイトにおける種子と苗捕食者による競争ととも組み合わせられる。 |
全体的には、低い植生密度のハビタットでの高い発芽と生存率はベイマツのコロニー形成が強く菌類の散布体に制限されていることを示すが、適地への到着と発生に成功した種の生存は土壌の窒素よりも初期遷移の競争に強く影響される。 [比較] Uesaka & Tsuyuzakiの論文は落葉樹のヤナギ属のパッチは特定の植物の植生侵入を促進する効果を持ち、常緑樹のシラタマノキ属のパッチは明らかな侵入抑制効果を持つという結果を示した。その理由として、落葉樹と常緑樹の葉のフェノロジー特性、幹・枝の構成の違いから光強度の抑制の影響が異なるためだと説明されていた。 一方、Titus & Bishopの論文では、若いルピナス・やせ地・日陰で土壌水分が多く競争が弱まったため苗の生存率が上昇した。植生密度の低いハビタットで高い発芽率と生存率が見られた事は、ベイマツの侵入が窒素固定細菌の散布体によって強く制限されていることと、適地への到着と発生に成功した種の生存は土壌の窒素よりも初期遷移の競争に強く影響されることを示した。 この二つの論文の違いは、Uesaka & Tsuyuzakiの方では先駆種が植生侵入を促進するが、Titus & Bishopでは先駆種がない場所の方が植生侵入が促進されるということである。しかし、ここで注目したいのは前者の論文が植物の種多様性で植生侵入をとらえているのに対し、後者では樹木であるベイマツの種子を植えてその発芽率や生存率で植生の定着度合いをとらえているということである。そもそもベイマツは、光条件が悪い(暗い)方が発芽しやすく、リターや下層植生が存在すると生存率が下がることが知られている(1)。Uesaka & Tsuyuzakiの論文では、ヤナギ属のパッチが適度な光強度と土壌水分を持っていたためそれらが直接的な要因として植生の定着を促進したが、Titus & Bishopの論文では弱い光強度、少ない下層植生、ルピナスの窒素固定細菌などが要因となって定着が促進されており、二つの論文の間では植生侵入の定義と促進要因が異なる。そのため、植生侵入の促進に関しては落葉樹・常緑樹間で比較する場合と、窒素固定細菌や光以外の条件に強く影響される場合を別にして論じた方がよいのではないかと考える。
|
自然環境学総論課題: 講義の内容は環境科学の最前線の話に特化すべきか、基礎的な生態学中心の話にすべきか。この講義は修士課程向けの講義ではあるが、3人の専門の異なる教官が3-5回ずつの講義を行う、という特性を持つため最前線のトピックを取り上げるという形が良いのではないか。 なぜならば、基礎的な内容と言っても、3?5回の講義で説明し終わることができないと思われためである。各教官はおそらく他講義で各専門の授業を基礎的な内容から掘り下げて説明していると思うので、基礎的内容はそちらで学ぶべきだと思う。 その一方でこの講義は、異なる専門の話が聞けるという長所がある。したがって、3つの視点から見た最前線のトピックを、修士の学生にも理解できるように講義していただければ、学生は広い視野で俯瞰的に現在の環境科学の流れを理解できる。この講義の形を活かした大きなメリットが生じるのではないかと私は考える。 生物圏修論発表会 修士1年生用 レポート2月12日(火)の発表で良いと思ったもの(1件)発表者: a) 他の発表と比べどの点が特に良いと思ったか |
研究により解明できたことが新しい発見であると感じられた点が良かったと思います。まだ発展途中の研究であるため、様々な実験を実行すればより内容が深まりそうで、今後にも期待できる点も良かったです。 b) 来年自分が発表するに際して、参考になった点 研究の意義を導入部分で述べていたが、結果らから得られた情報だけでは、その意義にまでつながっていなかったように思えます。発表の際には、導入→方法→結果→考察という流れを軸をぶらさずうったえる必要があると改めて認識しました。 2月13日(水)の発表で良いと思ったもの(1件) 発表者: a) 他の発表と比べどの点が特に良いと思ったか 非常に明瞭に研究結果が示されていました。他の発表と比べて得られた結果からこの考察に矛盾がありませんでした。また、情報量が適切で、かつ強弱をつけての発表だったので理解しやすかったです。 b) 来年自分が発表するに際して、参考になった点 示したいことを、得られた結果から素直に説明することが大事で、無理無理な結論づけをしなくて済むように実験・調査を組み立てる必要があると再確認できました。 必要以上に情報を出さず、シンプルにスライドをつくると分かりやすいと感じました。 |
エコパン大雨のため講義に切り替え、感想文を書いてもらった。攪乱についての講義の感想私は「撹乱と生物多様性-撹乱を利用する植物-」と題された講義を受けた。予定されていた植生調査の実習が行えなかった事は残念に思うが、講義で学んだ内容は実習との繋がりも感じさせ、とても興味深いものだったので満足している。本レポートでは、私が講義で学んだ内容から考えたことや調べたことをまとめる。噴火後の遷移の種類 私は高校生の時に、噴火が起こると群集は破壊され、その後一次遷移が始まる、と習った。しかし今回の講義で、挙げられた有珠山の例では、噴火後に火山 灰の下からいたどりイタドリなどの植物が成長していた。そこで私は、噴火後に起こる遷移には色々な種類があると考え、遷移の種類を左右する条件は何かについて調べてみた。 調査の結果、噴火後の遷移の種類を左右するのは火山噴出物の種類だという事が分かった。火山噴出物には溶岩、火砕物、火山ガスなどの種類があり、溶岩による撹乱では群集が壊滅するため一次遷移が起こるが、火山灰が薄く堆積した所や火山泥流が起きた所では群集が生き残り、二次遷移が起こる事がある。高校生の時に学んだ「噴火」とは溶岩を伴う噴火を指していたと考えられる。 |
群集をどの状態に保つか 講義で学んだ内容で最も印象に残ったのは、中規模撹乱仮説についての話であった。特に、生物多様性は中程度の撹乱の中で最も高くなるが、遷移の初期や極相にしか存在しない種がいる事を知って、群集をどの状態に保つのが良いのかという疑問を持った。近年自然保護が地球規模の課題としてよく挙げられるが、私は、どの状態の群集を保護するのかという基準を設ける事が重要だと考える。 遷移の初期や極相にしか存在しない種が存在する限り,生物多様性の高さを保つ事だけが重要だとは言えないので、私は、人間にとっての有用性を基準とする事が最も合理的だと思う。有用性とはその群集の中の生物が持つ物質などの価値の大きさである。ある範囲 毎にどの程度の撹乱があれば最も人間にとって有益な群集になるかを予測し、その状態を保つためにはどうすればよしいか考えることが、これからの自然保護にとって必要だと私は思った。 参考文献 [総説] 火山遷移初期動態に関する研究 (作成者:露崎史朗) (2012/07/19 訪問) |
エコパン植物が吸収するCO2量1) 実験目的木の周囲長や樹高から、その木の直径や体積などを求め、さらにその木が吸収した2酸化炭素量を推定する。また、その結果から、現在の“植林=エコ”という考え方を考察する。 2) 実験材料 メジャー、樹高計測器、樹高計測棒 3) 実験方法 ① 木の樹高と生枝下高を樹高計測器を使い測定した。周囲長をメジャーを使い計測した。 ② ①で得たデータを元に、木の直径、体積、重量、炭素量、吸収した2酸化炭素量を計算で求めた。なお、今回の木の体積は円柱で近似した。 4) 実験結果 測定データは以下のようであった。(表1省略) 表1より作成したデータは以下のようであった。(表2省略) 5) 考察 (1) 表2で求めた"炭素量"は最低値である。なぜか。 求めた炭素量は、最も太い幹に含まれる炭素の量である。よって、枝分かれした枝や、展開している葉、根などに含まれる炭素は結果に含まれていないと考えられる。だが、今回の測定時期は春であったため、葉の展開はあまり起きておらず、その分の誤差は少なかったと考えらあれる。また、落葉樹であるハルニレやシラカバなどは毎年、葉を落としているので、その分も炭素量が少なくなっていると考えられる。昆虫などによる食害や枯死量、植物自体の呼吸による炭素の排出も原因の1つと考えられる。 (2) 木が一年間に吸収するCO2はどれくらいか。 今回の測定した種類の中では、トドマツで樹齢が分かった、トドマツの樹齢は15年であったため、このトドマツの1年間に吸収したCO2量は平均で1.04 kgであると考えられる。 (3) "植林すること=エコ"という考え方は正しいのだろうか。 考察(2)より、トドマツの1年間に吸収するCO2量は1.04 kgであると推定された。日本人一人あたり1年間のCO2排出量は、JCCCA(全国地球温暖化防止活動センター)によると、9 t (2006年度)である。よってこのデータによると、日本人1人分のCO2を吸収させるためには、トドマツを8653本植えなければならないのである。このことを考えると百本や千本単位での植林は、CO2量の増加には、あまりにも微力で、エコにつながるとは考えられない。 しかし、植林することは悪いことではなく、自然環境の保全にはつながり、その点では"エコ"といえる。(もちろん、スギのみといった単一樹種のみの植林や、生息環境を無視した植林を除くが。)また、植林を通して、節電などを勧めることもできる。 これらのことから、"植林する"ことは、CO2量の増加にはほとんど意味をなさず"エコ"とはいえないが、自然保護や、それを通した省エネ活動の点では"エコ"といえると考えられる。 6) 感想 この実習を通して思ったことは、想像していたよりも、木が吸収したCO2量が少ないということだ。もちろん、CO2自体はとても小さい分子なので、数kg吸収した、というのはとてつもない量だが、それでも、もっと多くの量を吸収しているイメージがあった。特に考察(3)で、日本人一人当たり、9000本近くのトドマツがいるということには、とても驚いた。しかし、そのことにより、CMなどで流れる"当社は利益の何%を、植林事業に充てています"をもう一度考え直してみようと思った。 北大構内に生えている樹木の炭素量を測ってみよう1.調査目的今回の実習では、樹木1本ごとの炭素量を測定することを目的とする。実際のフィールドワークでは、面積当たりの炭素量を積算するため、プロットを作り、その面積内の全ての木の炭素量を測定するのだが、今回は省略する。また、樹齢の測定が困難であるため、考察時に概算でとらえる以外には、年間のCO2吸収量を計算することはしない。 2. 調査方法 北大構内に生えている樹木について、樹高、生枝下高(力枝まで高さ)、周長(根元より1.3 mの地点)を測定する。 |
3. 樹木1本当たりの炭素量の算出方法 樹高 H、生枝下高 h、周長L とする。樹木は根元から力枝までは円柱、力枝から最高点までは円錐と近似できる立体であるとする。すると、円柱部の半径は r= L/2πと表せるので、樹木体積Vは、V = (r × r × π × h) + {r × r × π(H - h)} × 1/3である。樹木の種ごとに異なる乾燥時の密度をρとおくと、樹木の重さWは、W = V × ρである。ここで、一般に、この重さの炭素割合は50パーセントなので、炭素量はW(C) = V × ρ × 1/2 となる。 4. 測定結果 実際に測定して概算した6種類の樹木の結果を以下に示す(表省略)。記号は上記3のものを用い、単位は H、h、L が [cm]、Vが [cm³]、ρ が [g/ cm³]、炭素量W(C)、CO2変換量W (C02)が[g]である。 5. 考察 5-1. 動物(ヒト)と植物(樹木)の個体数関係 この結果から、樹木により固定されたCO2の質量が分かる。地球環境センターの概算(※1)によれば、ヒトは 1日に約 1 kgのCO2を排出しているから、それぞれの樹木は、ヒト1人が1年間に排出する量として、およそ、ハルニ37年分、シラカバ2年分、イチイ半年分、セイヨウハコヤナギ12年分、ヒバ2年分、シダレヤナギ11年分のCO2を蓄えている計算になる。樹木の成長には少なくとも、この年数よりもかなり多くの年月がかかっていると考えられている(※ 2)ので、以下の 5-3で述べる測定値と実際の値の誤差を考えても、樹木は 1年間では、ヒトがその間に出す量のCO2でさえ、固定できないことになる。よって炭素循環が正常な生態系では、樹木は常に人間(動物)よりも多く存在することになる。これは食物連鎖における生物種間の量的関係(生態ピラミッド)と一致する。 5-2. 地球温暖化対策としての樹木利用 地球温暖化の原因とされるCO2の固定を期待される樹木だが、5-1でも述べたように、その炭素固定量は存外小さい。 5-3の誤差を補正(実測値を2倍)したうえで、今回測定した中で、炭素固定量の最も大きいハルニレの値を用いて計算しでも、ガソリン換算で5802 Lにすぎない(※ 3)。これだけ見ると多そうだが、石炭火力発電の消費電力で考えると、27634 kWhだけである。東京ドームで野球一試合4万 kWhであり、何十年もかけて固定した炭素量をたった数時間で越えてしまう。よって、地球温暖化対策として樹木によるCO2固定を活用しようと するならば、かなり大規模な森林としての保護、育成を行わなければ、ほとんど意味がない。 5-3. 実際のCO2素吸収量が実験値よりも大きい理由 実際に樹木が吸収するCO2量は、今回の実測値よりも大きいという。その理由は以下の3つが考えられる。 ・今回の実験では根の体積、質量を考慮していない。 ・同様に枝・葉の体積、質量を考慮していない。 ・吸収された炭素の一部は呼吸によって空気中に放出されてしまう。 特に根の大きさについては大きな誤差が生じる可能性がある。樹木において、地上部と地下部の乾燥質量はだいたい等しい(※ 5)ので、実際の値は、CO2固定量にして今回の結果の2倍近くになると考えられる。 6. 課題と展望 今回の測定により、樹木の炭素固定量がわかり、その誤差や限界、生態系の中での量的関係について言及することができた。しかし、樹齢がわからない弊害は大きい。個々の樹木が持つ炭素 量は、樹齢が異なるために単純比較することができず、種間の年間のCO2固定量の違いにまで論を発展させることができなかった。 しかし、もし種ごとの年間CO2固定量がわかれば、そして、それが種ごとに一定の値ならば、そこから樹齢を概算することもできる。炭素固定量の測定は、地球温暖化対策だけでなく、森林の管理にも利用できる可能性がある。 ※1 国立科学研究所 地球環境科学センター. ココが知りたい地球温暖化 ※2 今回調査したシラカバ、ヒバは樹齢30年を超えるとされる。 ※3 環境省「算定・報告・公表制度における算定方法・排出係数一覧」 ※4 「新エネルギー」がよくわかる本(早稲田聡(監修) P42-45, 56 (PHP文庫)参照し例示値を用いた。 ※5 佐藤孝夫. 1989. . 樹木の根の生長と分布特性. 光珠内季報 74: 8-12 |
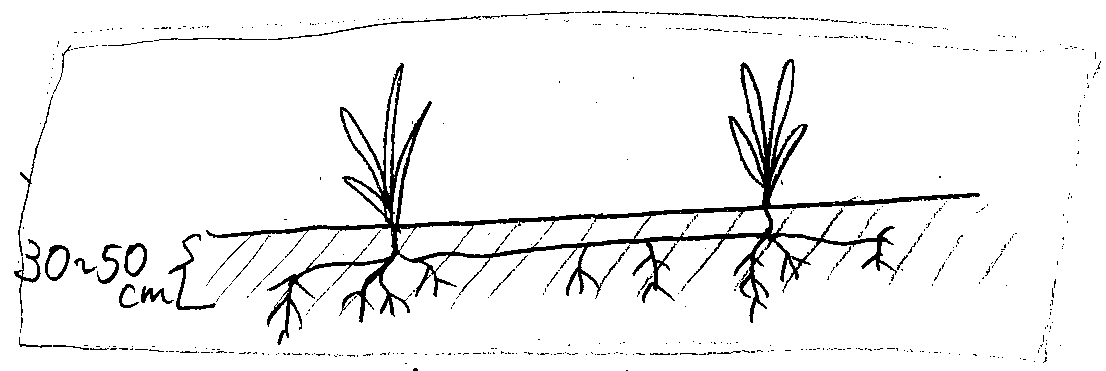
生命環境野外実習2010年10月21日。地下部スケッチのあとに30分程度で書いてもらったもの。大したもんです。[ 1 ] はじめに(目的)砂浜のような海辺は、植物が生息することが難しいと思われるが、今回の実習で行った久根別の砂浜にも何種類もの植物が生息していた。そのうち双子葉類と単子葉類(今回はウンランとコウボウムギ)を地上部も地下部も観ることによって、その場所に適応できている理由を探す。調査地と方法久根別小学校裏の砂浜で、スコップやシャベルを用いてウンランとコウボウムギを根から掘り起こして採集し、実験室に持ち帰る。持ち帰ったものをスケッチしながら観察する。結果と考察別紙にあるスケッチの通り、ウンランとゴボウムギは主根、側根を持つものと、ひげ根を持つものとの違いはあるものの、どちらも横方向へ広く根を伸ばすことによって、動きの多い砂浜でも安定して生息できると思われる。また、ウンランは、高さ15 cm程の低い花なので、風の強い浜辺でも花を咲かせやすいと思われる。また、スケッチした植物以外の植物も低いものが多かった。感想浜辺や砂浜で生息するには、根をしっかりと伸ばして、地下でしっかりと支えることがとても重要なのだと、自分で掘り起こすことによって、体感できました。また、他の特徴的な生息場所ではどのような植物が生息しているのか興味がわきました。[ 2 ] はじめに(目的)海辺の土壌は動きやすい。そのため通常の土壌に生えている植物とは違った工夫をもって生き抜かなければならないのだ。そこで今回、海辺に生える単子葉植物と双子葉植物を根こそぎ採取し、観察してスケッチすることにより、どのような生きる工夫をしているのかを探る。調査地と方法・調査地 → 久根別浜の海辺(七重浜付近)・方法 → 班ごとに指定された海辺植物をスコップやシャベルを使用して根が切れないように注意しながら採取する。 結果・地下部の形態双子葉植物も単子葉植物も地下茎が1 m以上あり、付近の同じ植物と連携して生えていた。また、地下茎は30~50 cm、もしくはそれ以上の深さまで根付いており、近辺の他の植物と根同士がからみ合っていた。 また、今回スケッチした、単子葉植物(コウボウシバ)、双子葉植物(ハマエンドウ)の地下茎の違いとしてハマエンドウの地下茎は一定の間隔でこぶのような節がみられた。また、掘っている際、ハマエンドウが非常に折れやすかった。
全体的に双子葉植物も単子葉植物も葉が厚くできていた。また、双子葉植物のハマボウフはテカテカした葉をもって低く生えていた。目立って高く存在したのはハマニンニクであり、その他は低く生えていた。 考察地上部の形態についてでは、葉がテカテカであったり、蒸散しにくい。厚く形成されているのは海辺の環境である「乾燥」から身を守るためである。また、低く生えているのは強風の吹きやすい環境に適応するためである。 |
地下部の形態については、地下茎や根が1 m以上あることから動きやすい土壌にも成育できるようにするためだと考えられる。それは、近辺の植物の根同士が互いにからみ合っている点からもいえる。また、地下茎や根が50 cmやそれ以上深く根付いていつのは、土壌の表面部分の方が動きやすいからではないだろうか。だから植物たちはより深く、奥へ奥へと根をのばし、安定した土壌に根をはるのだと考える。 ハマエンドウの地下茎に節があることについてはあまり自信がないが私の意見を述べる。 茎に節があるととても折れやすいと思う。そこで動く土壌に根付いているという点に注目した。もし、土壌が移動した際、そこにいる植物も動いてしまう。根が引っぱられたときに(移動で)茎が折れやすいと何か得なことがあるのではないかと思う。植物本体を支えることや、それ以外にも。だから節があるのではないだろうか。 感想根を切らないようにして植物を採取するのがとても難しかった。また、予想以上に根や地下茎が深くてとても驚いた。先生の話にも驚いた。(シロヨモギ)[ 3 ] はじめに(目的)植物は地上に出ている部分よりも、地下にはりめぐらされている根の部分の方が重要である。普段は見えていない根の部分がどのようになっているか、実際に掘って目で見ることを目的としている。また植物のはえている土地が海岸の砂浜であると、定期的に砂地が動いていることになるので、それにどう対応しているかも見る。双子葉植物と単子葉植物によっても根のつくりが異なるので、それについても見てみる。調査地と方法調査地 久根別浜の海岸沿い方法一班3~4人に分かれ、スコップとシャベル、透明な袋を用意する。海岸に群生する植物2~3種類(双子葉植物と単子葉植物一種ずつ以上)を、根を切らないように掘りかえして収集する。その際、根がどのように伸びているのかもよく見る。結果私たちの班は単子葉植物としてコウボウシバ、双子植物として白ヨモギを収集した。 コウボウシバはひげ根が細く短く部分的に密生していた。群生しており、横に伸びた茎でつながり、1つの群生で2-7に分岐しており、高さ5-15cm程度の高さに地上へ出ていた。群生するための地中の茎は1 m近くまで長いが、ひげ根自体はとても短かった。シロヨモギは主根が径0.5-1 cmで10 cm程度の枝のようになっており、そこから50 cm前後の細い側根が2,3本、短い側根が多数出ていた。根自体は浅く広く広がっていた。葉は裏に白い毛が密生しており、表面は丸みをおびていて、葉のふちは葉の裏側に反り返っていた。海岸全体で見ると、シロヨモギはところどころにまるくかたまって生えており、コウボウシバはコウボウムギに混じってポツポツと群生していた。 考察コウボウシバもシロヨモギも根はとても浅く生えていたため、砂地の移動では植物自体もそれに合わせて移動するという対応をしていると考えられる。また、どちらも海岸ではあまり沢山が砂浜を占めているような生え方をしていないため、海岸の植物の中では比較的劣勢で、強くはびこらない種類のように見える。砂浜の中でも、海の近くでは背の低い植物が多く、陸地に行くにしたがって、ススキやブタクサのように背の高い植物が増えてきていた。そこからみると、背の高いものは、高さの維持のために適度に安定した土地の方が適していると考えられる。そのため、移動しやすい砂地では、不安定な分背を低くし、広く根を広げることで対応していると考えられる。感想普段見られない根を見ることができ、また海辺という土地から、面白い根のタイプが見られて面白かった。コウボウシバもシロヨモギも小さな植物なのに、やたらと根が長くておどろきだった。植物の種類が変わる時の、土地や塩分などの違いが早く知りたい。高波ではどう対応するのかも気になって興味深かった。 |
生命環境野外実習目的: 東久根別海岸に生息している植物群落の植生調査を行い、単位面積あたりの植物の被度や種類、類似度を計測し解析することで、土壌移動攪乱と塩分ストレス勾配等が生じている海浜での植物相の環境適応様式や群落の構造の変化をグラフ化する等で視覚的に明らかにすることを目的とする。方法: 汀線から5 m程度離れた地点に杭を打ち、そこから内陸側へ巻尺を20 m程度伸ばし1 m四方の領域を巻尺と折尺を用いて作る。この1 m四方の領域の内の植物の種と被度をデータシートに記入していく。この作業を1 m-20 mまで繰り返し行い、データを集めていく。同様の工程を二度行うことで、二ヶ所のデータをサンプリングする。 準備するものは20 mの巻尺、杭、3本以上の折尺、データシート、1-2 mのコンベックスがあると高丈を計ることもできる。 結果 (別紙) 考察: グラフから考えられることを以下に述べていく。 1. 汀線から遠ざかるほど種数は増えていく。 「調査地1と調査地2の距離による種数の変化の比較」を見てわかるように、両地点ともに、杭からの距離が大きくなるほど種数が多くなる。また、この増加率も一定の増加ではなく、段階的な増加であり、両者に共通して、3-5 m, 6-7 m, 12-13 mでの増加が見られる。この地点では増加には、海水により塩分の影響が大きいと考えられる。特に12-13 mでの増加は高いため、この地点を境界に塩分ストレスに強い植物と弱い植物に分けることができると考えられる。またグラフから分かる事として、増加だけでなく、減少が必ず見られることが分かる。この減少は、海浜特有の植物相から、内陸の植物相への移行のキッカケと考えることもできるし、領域に植物がなかっただけとも考えられる。後者の場合、このことは海浜の植物は散在しても生息するものよりも、密生して生息するものが多いのではないかと考えることができる。 
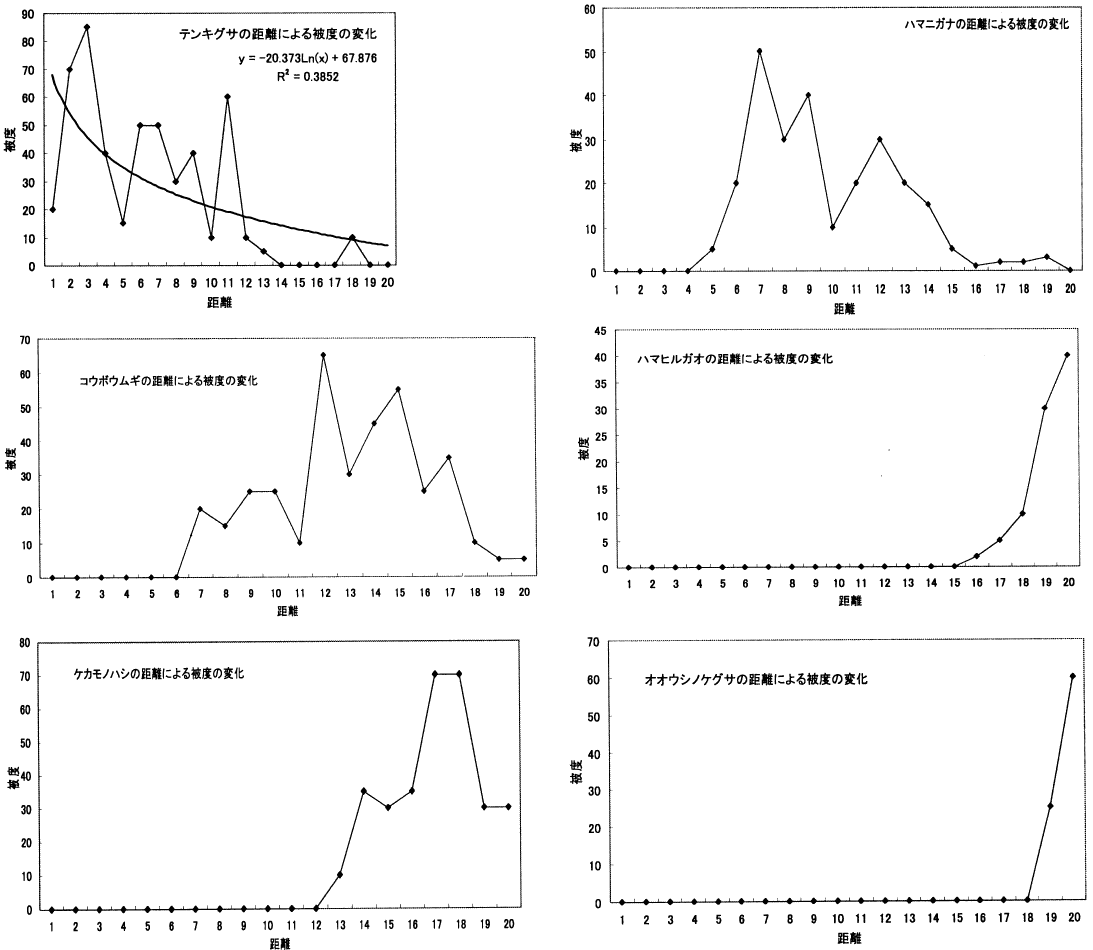
調査地1の距離による種数の変化_____調査地2の距離による種数の変化 2. 単子葉類の方が双子葉類よりも塩分ストレスに強い 「調査地1と調査地2の距離による被度の変化の比較」を見ると、領地点共に、テンキグサが海際から一番初めに生えていることが分かる。そして、後にコウボウムギが生え始め、ハマニガナが生え始めている。テンキグサもコウボウムギも多年生の単子葉類であることから。双子葉類よりも単子葉類のほうが塩分ストレスに強いと考えられる。(ちょっと極論) 3. 単子葉類は双子葉類よりも根の成長が早い。 海際は砂の移動が激しいと考えられる。このため太い主根と側根を発達、成長させる双子葉類は、根が土壌に定着するよりも早く移動による影響を受けてしまうため、成長することができず、一方、単子葉類は細いひげ根を多く成長させるため、表面積が双子葉類よりも大きくなるので、移動による影響を受けても耐えることができるので、グラフのようにテンキグサやコウボウムギなどが全体で見た時に早い地点で被度が高くなるのではないか。 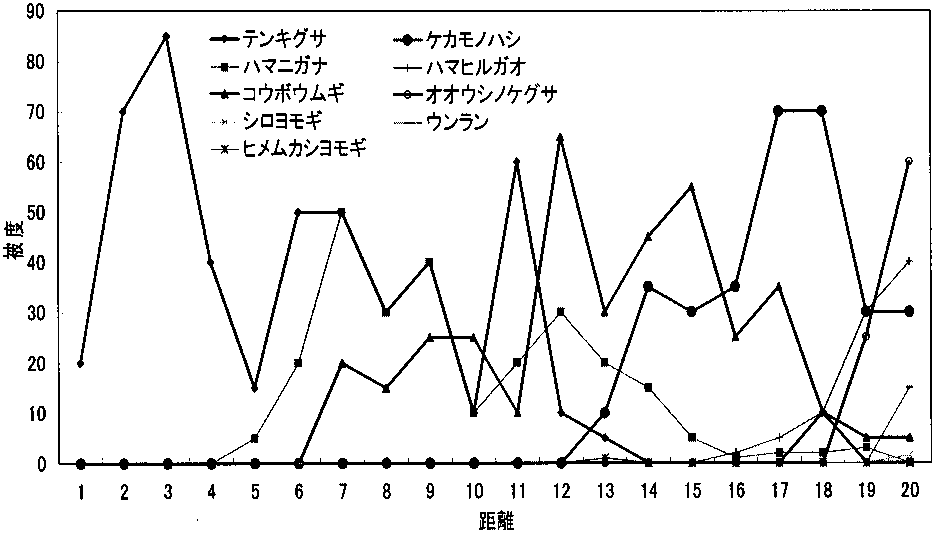
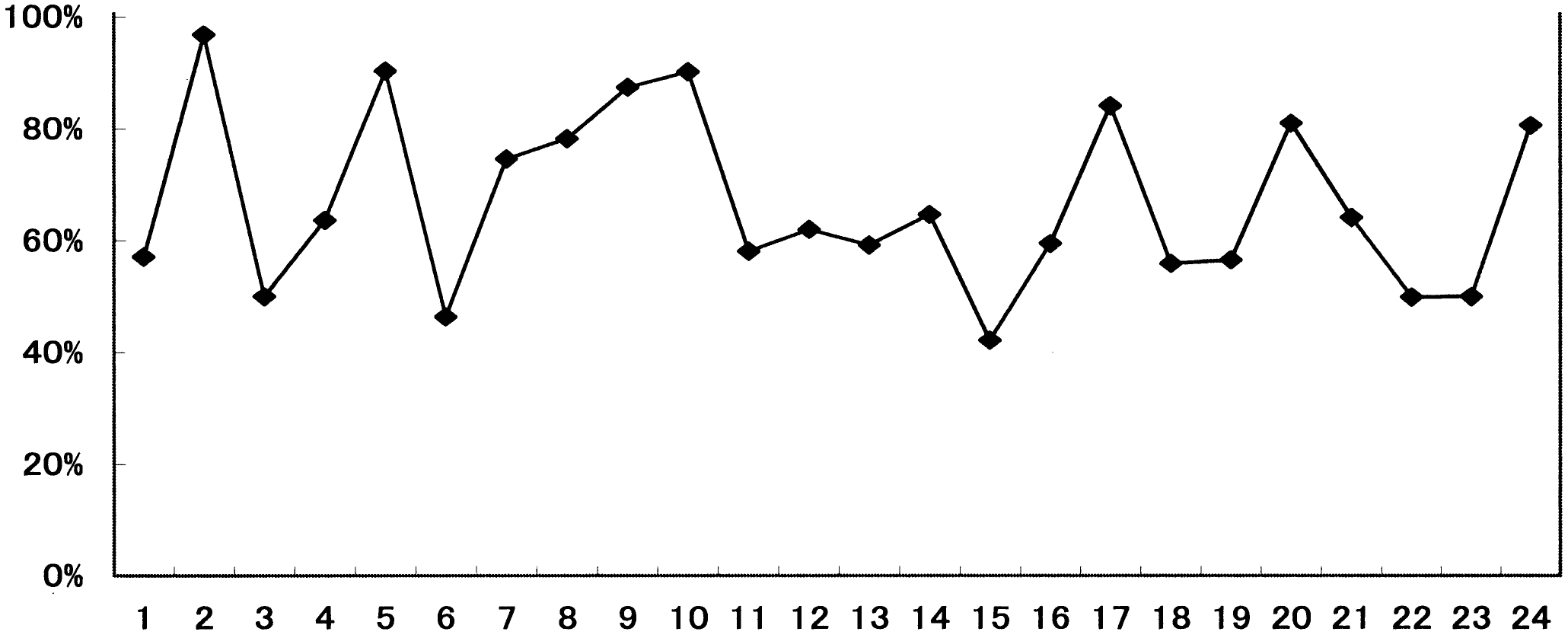
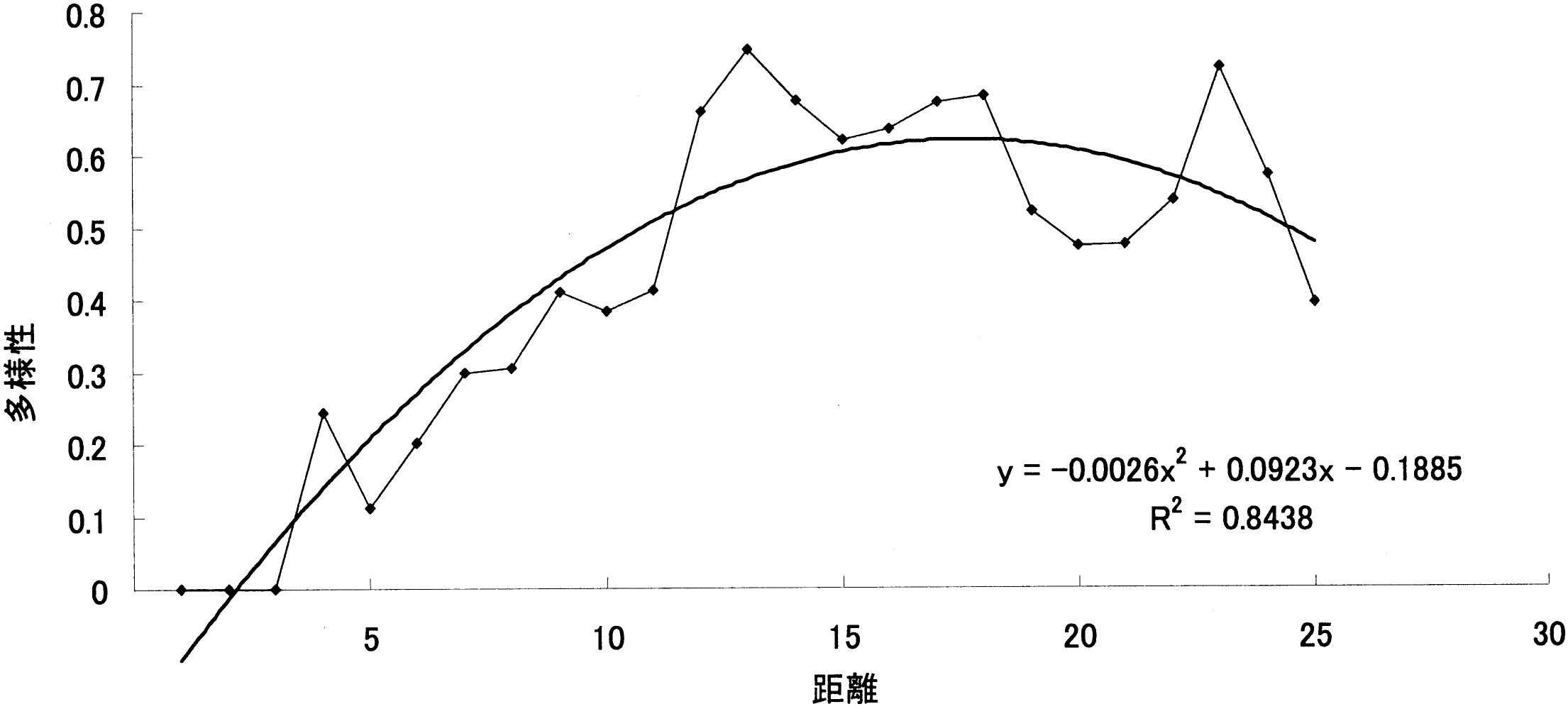
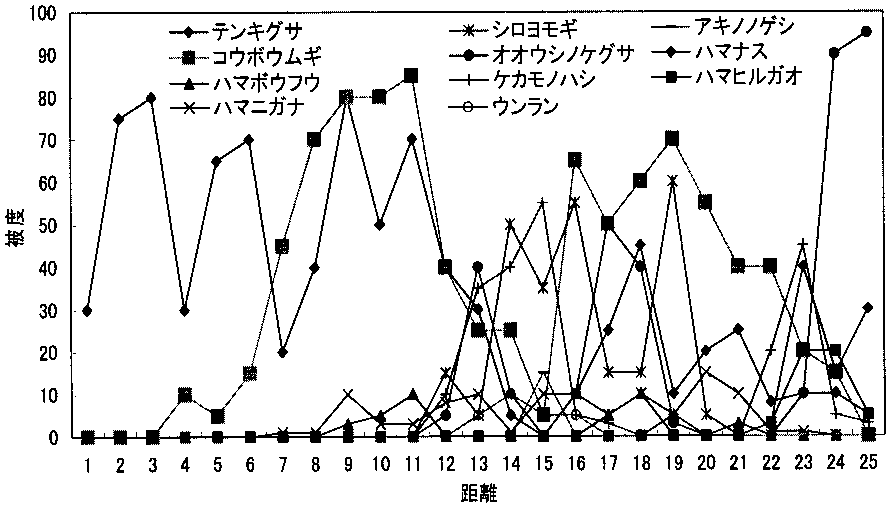 調査地1の距離による被度の変化_____調査地2の距離による被度の変化 4. 12 m以降で植物相の入れ替わりが起こる。 同様に「~被度の変化の比較」を見ると、12 m程度から双子葉類も増えており、一種よりむしろ全体の被度が多くなっている。また、植物相の入れ替わりもあり、この地点で土壌に何らかの変化があったことが考えられる。「ライン1」と「ライン2」から詳しい増減が確認できる。 5. 類似度は変化を繰り返す。 「調査地1と調査地2の類似度と距離の関係の比較」から分かる事として、類似度は一定にはならず、上昇と下降を繰り返している。また100%近くにはなるが100%にはならないと言える。上昇は、2つの領域の植物種のおのおのの被度が限りなく近くなることで起こる。一方、下降は新しい種の出現や古い種の減少・消滅がおこることで生じることがある。この事から、たとえ1 mでも植物相に違いが生じているのだと考える。 6. 全体的な種数が多くなると多様性は高くなる。 調査地1と調査地2は15 m地点で一種一種の被度はそれほど高くないが、多くの種がある。「~多様性の比較」のグラフを見ると多様性は高い位置にある。しかし、一種だけの被度が極端に高い所では、多様性は低い位置にある。このため、グラフから15-20 mの間が海浜では最も多様性に富んでいると考えられ、これは双子葉類が増えることに由来すると考えられる。このグラフは、この後のハマナスなどの被度が上がり減少し、その後、内陸性の植物が多く増えていくため多様性は上昇していくものと考えられる。 |
 ライン1 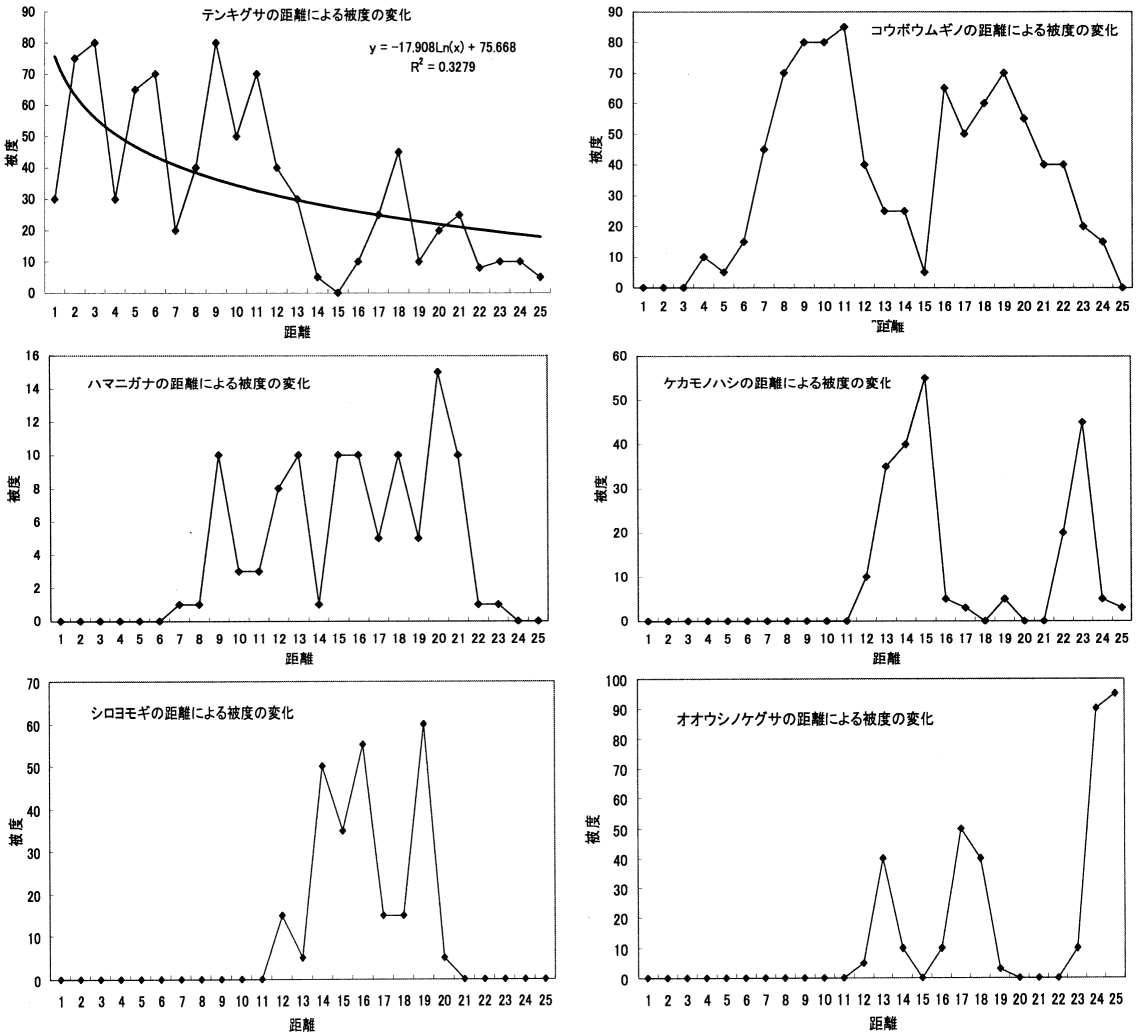 ライン2 
 調査地1の類似度と距離の関係_____調査地2の類似度と距離の関係 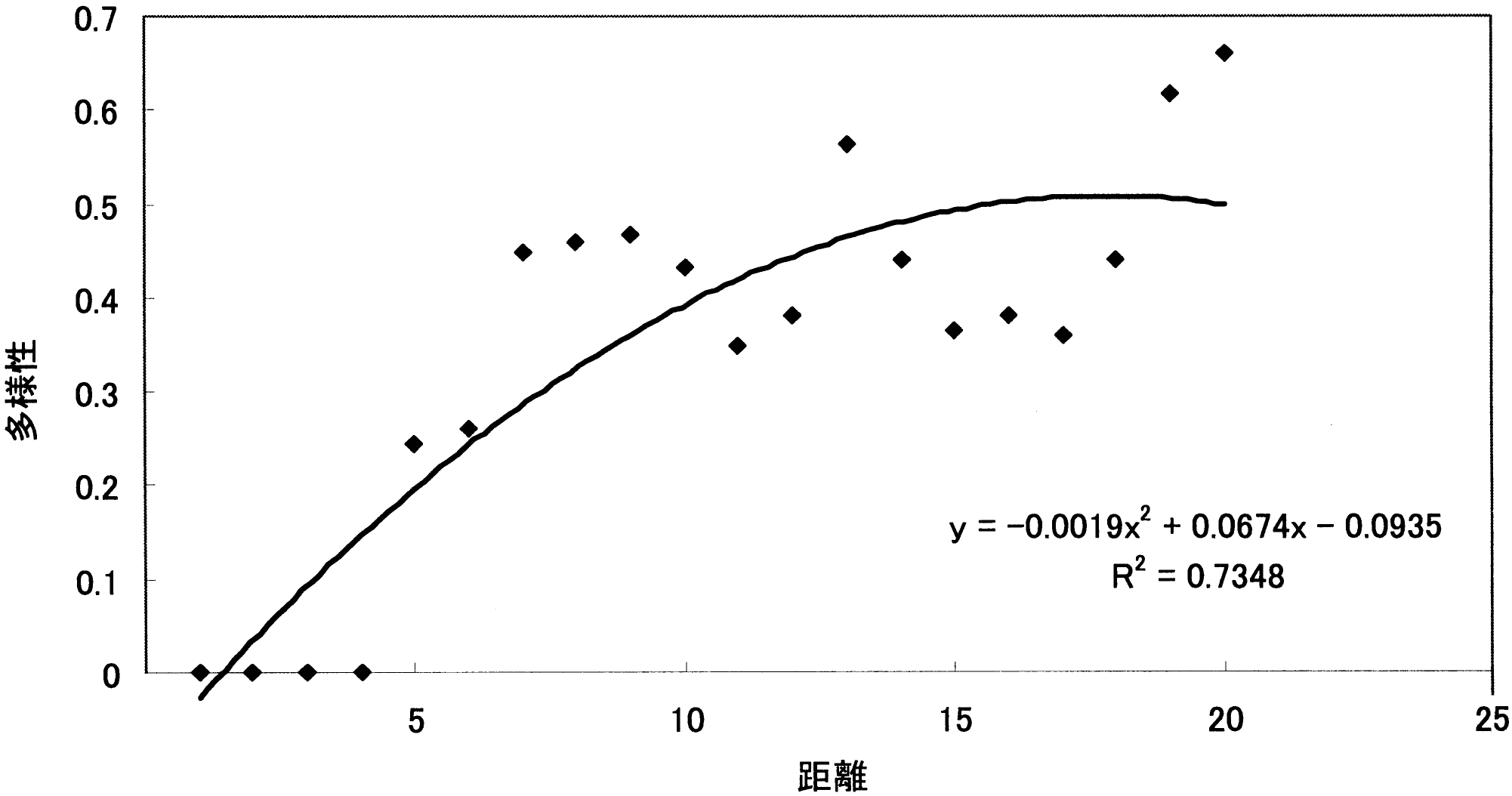
 調査地1の距離による多様性の変化___調査地2の距離による多様性の変化 |
環境保全学特論A問1. 選択した文献: 露崎史朗. 2001.1. 火山遷移は主要な自然撹乱の一つである。火山遷移初期植物群集の成立機構および動態は事象により様々であり、筆者は本稿で、これらについての先行研究を「植物に対する撹乱の度合いと頻度」「火山遷移の区分」「撹乱後の植物群集の動態」「帰化植物の侵入」「撹乱前の調査と永久調査区の必要性」という観点からレビューした。 この中の「撹乱後の植物群集の動態」では菌根と植物間の相互作用についても述べられている。すなわち、菌根の発達は土壌窒素・リンの吸収に影響するため植物群集構造に大きく関与することが指摘されてきた。しかしMSHでの菌根接種実験で、接種個体と非接種個体の成長に有意な差が認められなかったことから、菌根の存在が遷移上有効とする見解には疑問があるという。 2. Fujiyoshiら(2006)は、富士山の初期遷移段階に生育する4種の草本植物、Polygonum cuspidatum、Miscanthus oligostachyus、Aster ageratoides var. ovatus、Hedysarum vicioides、を用いて、AM菌の存在と土壌の発達がそれぞれ植物の成長に対してどのような影響があるかを調べた。 調査地で遷移初期の土壌(Bg)と遷移が進んだ土壌(Ic)を採取した。この土をポットに入れ、そこへ上記4種の現地性のAM菌接種個体と非接種個体を植えて実験室内で育て、その後乾燥重量、窒素含有量、リン含有量について計測した。 その結果、土壌はIcがBgに比べて4種の植物全ての成長成長を促進する傾向が見られた。またAM菌の接種は、H. vicioidesの成長を著しく促進したが、他の3種については有意な変化は見られず、その作用は種特異的なものである可能性が示唆された。 3. Fujiyoshiらは、菌根存在は植物成長に影響を与えることがあり、その影響は種特異的だろうという結論を出し、Titus ら(1998)は、菌根存在は植物成長に影響を与えないという結論を出した。両者は異なる結論を導いたが、両実験では用いた植物の種が異なる。Fujiyoshiらは4種の植物を用いた。このうちマメ科のH. vicioidesは窒素固定を行う種で、実際本種がAM菌の接種により著しく成長が促進された。一方Titusら(1998)は7種の植物を用いたが、窒素固定を行う種は含まれていない。また室内で行ったFujiyoshiらの実験に対し、Titusら(1998)は屋外実験で、コンタミの可能性が否定できないのはTitus本人も認めるところである。 4. 露崎(2001)は「菌根の存在が遷移上有効である」という見解に対し、Titusら(1998)の結論を引用して、この「再検討が必要である」としている。しかし上述した4のように、菌根の存在が遷移上有効であるかという問いに対しては、実験に用いる種、つまり現地に生育する種に窒素固定を行うものが存在するかどうかが深く関わってくる。このように菌根と初期遷移の関係は一概に言えないことから、本文は「菌根の存在が遷移上有効かどうかという問いに対してはTitusら(1998)やFujiyoshiらなど対立する見解があり、さらなる事例の蓄積が必要である」と述べるのが妥当であろう。 |
B1. 生態系から放出されるメタンについてまとめる。地球温暖化による生態系からのメタン放出増大には、大きく分けて2つあると考えられてきた。1つはツンドラ地帯の永久凍土に含まれるメタンの排出がある。これは温暖化により永久凍土が溶け、その結果として放出されるものである。2つめは湿原地帯から放出されるメタンである。これは湿地に生息するメタン生成菌である嫌気性細菌が温暖化によってそれの活動が活発となり、メタン放出が増大するものである。しかし、陸上植物からも大量にメタン放出があることが発見された(Keppler et al. 2006)。これにより、森林化による植生の増加によっての地球温暖化緩和に対し、疑問が出てきた。2. Parsons et al. (2006) は、Keppler et al. が実験室の結果を地球規模にスケールアップする際に、問題があると指摘した。これは茎や根を含む純一次生産速度(NPP)を用いたことに、メタン放出と植物の成長に関係があるという証拠が無く、葉と同じレベルで茎や根からメタンが発生するとは考えにくいためである。そのためParsons et al.の研究では、NPPの代わりに葉の現存量をスケーリングに使って推定した。その結果、陸上植物からのメタン放出量はKeppler et al.の推定量よりも全体で約72%少ないとわかった。 3. Parsons et al.とKeppler et al.では、全球での各バイオーム推定量に大きな違いが見られる。問題2のところでも書いたように、Parsons et al.では全体のメタン放出量はKeppler et al.の推定量より約72%少ない結果となったが、さらに熱帯雨林帯での放出量は-80%、温帯の森林地帯で-55%、熱帯のサバンナや草原地帯でも-73%と Parsons et al.でのメタン放出量の見積もりは、かなり小さいものであると結論づけている。 4. 以上のことから、Keppler et al.で発見された、陸上植物からメタンが発生しているという事実は、Parsons et al.の研究でも確認されたが、両者での研究が示めしたメタン発生量の見積もりには大きな差があり、どの程度メタンが発生しているのかは、未だ不確実なことが多い。そのため、温暖化物質としてとても重要であり、地球温暖化に大きく関与していると考えられる大気中のメタンがどこから発生しているかを解明のために、陸上植物からの全球規模でのメタン発生量をさらに正確に、定量的に示す必要があると考えられる。 参考文献
|
景観生態学レポートTheme 1: α-diversityα-diversity (= intra-community diversity)とは生物多様性に関する概念の1つで、ある均質な生育地にどれだけの種が共存できるかという群集内多様性の尺度として用いられる.つまりα-diversity とは、局所的な種の多様性を示す.したがって地域間でα-diversity を比較する場合は、その地域に卓越する中性的な環境に成立する群集を選んで種数を比較する手法が使われる.それに対してβ-diversity(= inter-community diversity)とは、生息地の多様性に基づいた生物多様性の概念であり、各々の立地に対応した群集型や植生型の数によって表されることが多い. おおまかな傾向として種の多様性は対象面積を広げるにしたがって増加する、これは対象面積自体の拡大によって生息地が多様化し、出現種数が増加するためと解釈できる(β-diversityの増大). その一方で、対象面積の拡大による種多様性の増加は、β-diversity のみならずα-diversity にも影響を与えると考えられる.α-diversity は、局所的な生息地における種間競争や共存の結果を反映するだけでなく、種の供給源としての広域的な種の多様性の影響下にあるからである.広域的生物相が潜在的な種の供給源としてのプールとなり、α-diversity に作用するのかもしれない.景観レベルでの広域的な種多様性はγ-diversity (landscape diversity)と呼ばれる. Theme 2: Relationship between α-diversity and γ-diversity Patel et al. (1996)は東欧エストニア(面積4500km²)を対象として、植生学的に広域的な種プールと局所的な種のプールおよび小プロット(1 m²)に出現する平均植物種数の関係を調査した.彼らは広域的プールの定義を、気候・自然地理的に均一であり様々な群集の共存が可能な地域における種のプールとし、広域的プールの調査範囲をエストニアと定めた.一方、局所的プールは、ある1つの植生景観で占められる群集内において共存可能な種のプールと定め、14の植生タイプを局所的プールの範囲と見なした.その結果は、広域的プールと局所的プールの間には正の相関が認められ、更に局所的プールと1 m²プロットの出現種数の間にも同様の関係が認められた.各スケールにおいて供給源としてのプールと種の多様性には比例関係があり、種の多様性はスケールによって飽和せずに種プールによって規定されているのかも知れないと結論づけた. 一般に植物社会学では、植生タイプの階層分けに様々な解釈が可能であり、群集タイプの増加を客観的に計測することは難しい.Patel et al.は、植生タイプを決定するためにエストニアにおいて観察された維管束植物1416 種の中から1073種を選び出しているが、その判断基準をEllenberg らによる欧州中部の植生指数に求めている.そのためEllenberg のリストに存在しない種については考慮されていない.このように、植生タイプと対応するように対象植物種を選択しているが、判断基準は提示されていない(植物種の抽出により現実の種プール値よりも算出プール値が小さくことに関しては補正されている). |
Patel et al.の論文は、ある地域の植物群に関する解析例だが、地球上の様々な生物群における解析例も報告されている.Caley and Schluter (1997)は、各大陸において、異なった生物群(脊椎動物、無脊椎動物、植物)を取り上げ、局所的多様性と広域的多様性との対応関係は検討した.
|
生態系構造学地表面の変動と遷移系列における地衣・コケ類の侵入との関係1. はじめに(レポートにまとめた部分)"火山遷移は一次遷移か"(露崎,1993)では、火山性の一次遷移系列において地衣・コケ期が存在する必然性がないこと、逆に"A rolling stone gathers no moss"の言葉のように、地表面が撹乱されている環境では、地衣類やコケ類は侵入しにくいことが指摘されている。さらに、土壌の肥沃化、富栄養化に関して、これらの植物群が存在しなくても、維管束植物は定着できることが示されている。ここでは、遷移系列において特に先駆的と誤解されがちな地衣類を主に、その土壌形成作用を考えるとともに、課題論文中で話題になっている地表面の撹乱や変動と、遷移に伴う植物群落の形成とを、等価で捉え直し、より多角的なバイオスフィアの時間的変化の概念を示す。 2. 参照した文献の内容遷移初期の地衣類の侵入過程については、火山よりも欧州や北米の氷河後退による裸地で多く観察されてきた経緯がある。ここでは地衣類は、その耐乾性と貧栄養環境につりあった低い成長率、岩石中にも菌糸を伸ばす定着力のおかげで、他の植物とほとんど競争せずに岩石上に侵入する。そして雨水を吸収し、その中にわずかに含まれる窒素成分を蓄積したり、岩石中の成分を少しずつ抽出して、同化作用を営む。また、岩石上に出現するStereocaulon属やPeltigera属では、窒素固定のためのシアノバクテリアを共生させる種も多く、マメ科植物と同様の先駆性を持つ。そしてこれらの地衣体の侵入により、岩石表面は吸水と乾燥を繰り返し、weathered rind (タマネギ状風化の皮)を生じると考えられる。さらに、地衣体が放出する各種の地衣成分(有機酸)は岩石の化学的風化を促進する。 他方、地衣類には多様なニッチを持つものが存在し、その種多様性は維管束植物と同様に、むしろsubclimaxな群集で高くなる。温帯の森林が極相に至る時、地衣類群集もそれにあわせて極相化し、Lobaria属やRamalina属などが特徴的となる。遷移初期に出現する種は、地衣類の中でもweedy (荒れ地戦略)な種であり、多数の小さな胞子を生産する(以上Topham, 1977)。 次に、地衣類侵入の基物となる岩石や土壌の安定性について考えると、地表面は常に水や風の流れによって侵食作用(erosion)を受けており、これらの粒子は運搬される傾向がある。また前述の風化作用によって形成された地表面の粒子も、重力や凍結融解によって多様な移動(mass movement)をおこす。これらの動きは、小さな構造土の形成から火山性の泥流による巨石の運搬、地層の隆起・沈降に至るまで、いろいろなスケールで、しかもいろいろな時間軸で起こっている。特に、本論で問題となっている安山岩性の火山噴出物の堆積した地表面では、これらの移動は大規模で速く、植物の定着に影響を与える(東, 1979)。 3. 1. 課題論文と2. 文献の内容の相違点地表面の変動が地衣・コケ期の存否に影響を与えているという現象においては矛盾点は見られないが、その後のいくつかの分析や遷移の分類に相違点が見られる。 まず1.では、地衣・コケ期の存否を一次遷移の特徴ととらえており、それに疑問を投げかけているのに対して、2.では特に地衣類に関して、遷移初期の岩石や土壌に出現しやすい種と後から侵入してくる種が存在することを述べていて、種によって遷移系列中に占めるニッチが異なることを示している。 |
さらに1.では、地衣類とコケ類による土壌形成作用を、土壌の肥沃化・富栄養化の過程と位置付け、窒素固定能力を持つことの重要性を述べているのに対し、2.では、それ以前の岩石の機械的・化学的風化の過程を問題にし、地衣類が付着した場合、どのようにして安定な岩体が細粒化されて維管束植物の侵入に適した土壌を形成するのかを述べている。 また1.では、土壌や岩体が不安定な場合には、教科書的な一次遷移系列から地衣・コケ期を除くべきであると指摘しているのに対し、2.では地表面の変動をいろいろなスケールで主体的に捉えており、氷河の後退地、いろいろな性質を持つマグマによる火山活動、侵食や移動によって多様な裸地が出現し、その各々に適した先駆植物が侵入してくることを示唆している。 4. より多角的な遷移の概念遷移初期の岩石の風化や土壌の形成パターンは多様であり、例えば岩体が安定で表面が乾燥し貧栄養的な場所では、地衣類のうち先駆的な種が侵入し、風化の結果もたらされた岩石の裂け目には、その環境に適したCladonia属やコケ類などが侵入する。しかし、それと隣接する場所に侵入する植物群落は、その場所の地表面の撹乱の度合によって全く異なるものになることから、植物が次々と環境を変えていく(ecological succession)のと少なくとも等価な概念で地表面が変動していく(geological succession)と考えるほうが無理がない。 私見になるが、十勝岳北西斜面のフィールドでは、地衣類の侵入は火山噴出物の堆積年代よりもその後の地表面の移動に大きく左右されており、細かい火山砕屑物の崩れやすい斜面には地衣類は無く、地表に露出した溶岩や泥流によって下部に移動してきた巨岩には、その岩質に応じた特徴的な地衣種の侵入が見られる。また、周氷河地形の構造土についても、多角形の各辺に集合して安定化した大きなレキには多様な地衣種が付着しているが、多角形中央部の回転や移動をおこしやすい小レキ群には地衣類の侵入はほとんど無い。 以上のことから、地表面はマクロからミクロなスケールまで常に何らかの形で揺り動かされ、その中に色々な戦略を持った植物が入り込んでくると考えたい。遷移初期が往々にして貧栄養的で乾燥した環境であったため、地衣類やコケ類などの存在が目についたものと理解している。このように考えていくと、課題論文の最後の表現のように、一次遷移と二次遷移を区別する以前に考えるべき問題が多く存在することがわかる。長く理科教育にかかわってきた立場から言えば、自省の念とともにより正確な内容を新しい世代に伝える責任があると感じている。 文献
|
|
伊豆中央高校から感想文が送られてきた。嬉しい限りだけど、自分だけが持ってるのはもったいないので、無記名なので公開することとしました。原文は、手書きで、その方が味があっていいのですが、ワープロで打ち直してます。また、誤字・脱字とかも、そのままにしてあります。 先生からの手紙 (個人名は隠す) 平成21年11月18日
北海道大学
静岡県立伊豆中央高等学校 出前講義について (お礼)
寒気の候、ますます御清栄のこととお喜び申し上げます。 |
|
|
専門的な用語がたくさん出てきていたけど、スクリーンの絵や図などでとてもわかりやすく表現されていて理解しやすかった。
今回の話は、生物だけでなく地学なども合わせて考えるような話でした。パソコンで、森林の図や、コケの分布のグラフなどを用意してありとても見やすく分かりやすかったです。 僕は今日の講義を受けて地球温暖化という問題ついて新しい印象を持ちました。僕は生物を選択しておらず、植物に関してはあまり興味がありませんでした。この講義を受けようと思ったのはどちらかというと地球温暖化に対して関心があったからです。今までも、地球温暖化については自分で調べたり、知識を集めたりしていたので、今日の講義にあった生態系変化予測などは見たことがありました。しかし、植物が実際には予測通りにいかないことは今日の講義で初めて知ることができました。地球温暖化という大きな問題を解決するために、多くのことを学ぶ必要があるということが分かりました。これからは一つの方面からではなく、様々な学問・分野から地球温暖化について調べたいと思います。そして、学んだ情報に対して常に疑うことを忘れずにいきたいです。 今回の講義は、環境の変化と植物の関係についてという、日常では気にならないことだったため、初めて知ったことばかりであった。まず、有珠山における火山噴火後の植物の変化では、苔類、地衣類の進入が最初ということをくつがえす結果となり、とても驚きました。また、森林火災後でのクロトウヒの成長は、年々地表が燃えることにより、少なくなっていた。それだけでなく、広葉樹の成長が多くなっていることに驚きました。こうした環境の変化によった、植物の変化の研究をすることで、環境保全に役立つことが分かり、是非これからも研究を続けてほしいと思います。自分も物事を疑い、興味を持ち、研究し、理解することをしていきたいと思いました。 今日の講義は、生物にも興味があって、北海道大学も行きたい大学だったので、聞いて、とてもおもしろかった。すごい研究をしていると思って聞いて、山火事から、何年後かたった後、さらに何年後かたった後に研究して、そこから、その時期に、存在する植物の種類を調べて、その結果から、さらにまた調べるということが、おもしろいなと思った。 特に、最後の、地球温暖化で、植物が生き残るためには、動物と違い、動けないので、種をとばさないと生き残れないけれども、植物の種のとばす距離は、そこにたりず、生き残れるか?ということがすごく興味を持ちました。あと、それをどういう風に解決していくのか・・・ということを自分で調べたいと思いました。 攪乱があった時最初に苔類から一年生草本、多年生草本が生えてくると聞いてそうだろうと思ったけど火山灰では動くから苔は生えず、一年生草本は存在しないから生えないということを聞いてびっくりしました。そして多年生草本の生えたところからその栄養で森ができるのかと思ったらまったくちがうところにできるということを聞いてまたびっくりしました。クロトウヒのことは焼けてしまうことを防いでいかないと絶滅してしまうそうなのでなんとかして生態系を保っていってほしいです。温暖化での生態系の移住はどうなってしまうのかとても興味があります。これからは生物学にもっと興味をもっていきたいです。
僕は、この講義のテーマが「環境」ということで、正直なところどのような内容なのかまったくわからなかったけど、講義を聞いて、今現在、「環境」に関してあまり良くないということがわかりました。 |
今日は「錯乱と空き地と生態遷移」というテーマで、講義していただきました。この講義で一番印象に残っているのは、桜島の溶岩の上では、400年たっても林にならないということです。僕は100年くらいで森林まで回復すると思っていたのでとても驚きでした。
他にも温暖化に伴った生態系の変化予測で植物が移住できる範囲を予測したり、もし、移住できなかった場合は、その種は消滅してしまうのか、それとも移住できなかったとしても、また新しい生態系がその場で生まれてくるのか、といった、どんどんと様々な疑問、想像が広がり、生まれてきました。アインシュタイン、カール・マルクスが言ったとおり、疑問を持つということは、とても素晴らしいことなんだと改めて思いました。 地球環境科学なんていう難しそうな名前がついていたので、講義が分かるだろうか、と不安な面もありました。しかし、けっこう分かりやすかったです。環境科学おもしろいものだなあと、親近感がわきました。私は勉強はそんなにはできませんが、もっとがんばっていきたいと思いました。 今日の講義を聞いて、生態学部はどのような研究をしているのか、ということと、いかにあらゆることを疑問にもつことが大切なことかが自分なりに分かりました。露崎先生の今の生活があるのも疑問をもつことから始まっていて、そのことから教科書のことが完全に当てはまっているわけではないことが、30年以上も研究を続けて、変遷のグラフを年代ごとに書きつづけていたりと、多大な成果を挙げられていることも分かりました。そして、その中からもさまざまな角度からの疑問でさらに研究を続けていって、近い将来には大いに役立っていくかもしれないということも知れて、露崎先生は偉大な方だなと思いました。自身の疑問から次々と展開されていって他者にもその影響が来るかもしれないということはすごいことだと思います。僕は、科学分野に進んでいくのかはまだ分かりませんが、僕も科学でないことにもどんどん興味を持って物事を考えていけるようにしたいと思いました。 教科書に違うことが実際に起きていることに驚きました。植物はいろいろな攪乱によってたくさん死んでしまうけれど土の中で生き残った種や別の場所からやって来た植物などによって空き地が再び森に変わっていくということを聞いて、植物の生命力のすごさを感じました。また、植物にもいろいろな種類があって攪乱に強いものや弱いもの、成長が早いもの遅いもの、栄養がたくさん必要なものあまり必要でないものなど、それらの違いによってその地域でどれだけ繁栄するのかがすごくよくわかりました。そして、マルクスとアインシュタインの言葉がすごくよかったです。
今日、北海道大学の方に、生態関係について教えていただきました。講義をして下さった方は、いつも「○○だとどうなるか?」「□□とすると、△△はどうなっていくのか?」など、疑問をいつも一つ抱えながら説明してくださったのでとても解りやすかったです。
今日、この講義を受けて、とても地球環境や、植物群集について、興味を持てました。すごく分かりやすかったので、スムーズに理解でき、「どうしてだろう」「なんでだろう」と思うことができました。
分かりやすい講義ありがとうございました。
今日の講義で生物のことについて関心をもつことができた。 |
地球温暖化生態学特論レポート環境科学院 環境起学専攻 修士1年 76103020 山田翔一 生態系の変動を予測する情報を得るにはその生態系の規模を考慮する必要がある。なぜなら、生物の分布要因は規模によって変わるからである。その生態系の区分方法には規模によってさまざまなものがある。そのなかでも最大の規模つまり地球規模での生態系区分はバイオームと呼ばれる。バイオームの区分要因は温度と降水量である。生態系の分布には先ほどまでの空間規模のほかに時間規模も考える必要がある。空間規模と時間規模には対応関係がある。時間軸に対する植物の応答は大きく2つに分けられる。1つは短期応答である。これは短い期間で起こる反応であり、光合成速度や呼吸速度などの変化は環境変化に対してすぐに応答した短期応答である。そして、それらの累積した結果である生産力変化や生態系の移動などは長期応答になる。つまり、短期的な変化が起こってもそれが累積されない限り、長期的な変化にはならないということである。 生態系の変動を知るには、その生態系の構造と機能を空間規模と時間軸で調べる必要がある。その生態系が小規模であれば現地での追跡調査を行えるが、大規模になるにつれ現地での追跡調査が難しくなっていく。そのような大規模での調査で必要となるものがリモートセンシングである。そして、リモートセンシングで得られたデータの解析にはGISがよく使われている。 植物の葉は光合成の際によく吸収する波長とよく反射される波長がある。植物の多い所ではよく吸収される波長とよく反射される波長の吸収率の比や差が大きくなるので、リモートセンシングではその原理を利用して地上を覆う植物量を推定する。そのすいて似も通られるのが植性指数であり、リモートセンシングにおけるもっとも基礎的なデータである。 生態系が温暖化の影響を受けることによって、その分布パターンが変化すると、その生態系が放出あるいは吸収するCO2量をはじめとする物質循環量が変化する。生態系が吸収するCO2量が減少すれば、いままで吸収できていた分のCO2分は大気中に残り、温暖化は加速されることになる。このように環境変動が生態系を変え、その生態系が環境を変えるようなフィードバックが存在する。このようなフィードバックは多くの生態系の変化を見る上で重要である。 "第6章 地球温暖化にともなう陸上生態系の変化"を読み生態系の "規模"について詳しく知ることができました。規模には時間と空間の2つがあり、規模の大きさを考えることによって同じ生態系でも異なる事実が判明し、生態系を理解するには規模というものを考えることがとても大切であることが分かりました。 |
環境保全学特論レポート環境起学専攻 76103020 山田翔一 遷移とは植物群落が時間とともに一定の方向に変化していくことである。最初の状態によって一次遷移と二次遷移に分かられ、一次遷移はされに乾性遷移と湿性遷移に分けられる。一次遷移とは火山噴火で生じた溶岩台地や新たに出現した島、大きく浸食された崖などのように植物が存在しない裸地から始まる遷移である。新たに露出した裸地での遷移は、その一連の変化が完了するまでに数百年要する。二次遷移とは森林を伐採した跡地や山火事が起こった跡地のように、今まであった群落が不完全に破壊されたところから始まる遷移である。一次遷移とは異なり、すでに土壌が形成されており、埋土種子や地下部が残っているため一次遷移に比べて速く進行する。乾性遷移とは乾燥地から始まる一次遷移のことであり、湿性遷移とは湖や湖沼から始まる一次遷移のことである。遷移初期段階の種は生息場所の条件や利用できる資源の量を変化させ、新たな種の侵入を可能にする。この過程を促進と呼ぶ。この過程は、初期の条件が厳しい一次遷移では非常に重要である。遷移の後期に侵入してくる樹木は遷移初期の種と遷移後期の朱に区分できる。遷移初期の種の特料は、葉群が多層構造になっている点である。葉は樹冠から補償点を越える光が受けられる深さの内部まで生い茂る。よってそのような種は遷移初期段階で豊富に利用出来る光を効率よく得ることができる。それとは対照的に、単層の葉群の種が存在し、それらの種は木の周囲に単層の葉群をまとい、遷移後期に密度が高くなった樹冠で効率よく光を獲得できる。もし、多層構造と単層構造の葉群をもつ種ができたての空所に同時に定着した場合、普段は多層構造の種が速く育ち、となりの個体と接触するようになるまではそこで優占するが、さらに混み合ってくると単層構造を持つ種が優占するようになる。遷移は進行すると安定した状態に落ち着く、この状態を極相と呼び、その状態に達した群落を極相群落と呼ぶ。極相の概念については長い歴史がある。初期の段階ではFrederic Clements(1916)によって、それぞれの気候帯には、すべての遷移の最終段階としてたった一つの極相があるという考え(単一極相)が提唱された。この単一極相の考えには多くの生態学者が意を唱え、その代表の一人であるTansley(1939)は多極相という考えを提唱した。この考えは1つの気候帯でもその中には、その地域の気候、土壌条件、地形、野火などの要因の組み合わせによって様々な形の極相が存在するというものである。さらにその後、Whitaker(1953)が極相パターン仮説を提唱した。この仮説では、極相は環境傾度に沿って変化してゆく連続体であり、独立した極相として分離することはできない。 |