(2024年4月24日更新) [ 日本語 | English ]
| 日本語 | 英語 |
|---|---|
|
ここからは「学生の業績報告」だかのためのメモ
|
発表をテープで起して若干手をいれたもの(さらに手直し)
旭川市大雪クリスタルホール大会議室
1999年3月20日 13:00-17:00
|
北大院地球環境の露崎です。馬渡先生の紹介にありましたように, 私は1980年北大に入学し卒論研究で有珠山の植生回復に関する研究を始め, それから15年が経過しました。この間, ずっと有珠山で毎年毎年同じ所に行って何が生えているかを記録し, 火山の噴火後, 植物がこういうふうに移り変わってきている, そしてその理由はなんなのだろうかという研究をしてきました。北海道は火山島で, ここの近くには十勝岳というA級火山もありますし, 比較的身近な話になるものと思い, その研究紹介をさせて頂きます。 まず遷移とはどのようなイメージなのかですが, その一般的な考え方を図1に示しました。X軸に時間をとり, その時間の最初の頃, この場合では左端が, 例えば火山の噴火直後, あるいは山火事直後であるとし, そのようなある撹乱によってそこの植物が全くあるいはほとんどなくなってしまった時を0年とします。右端は, 10年のこともあれば100年のこともあれば1000年のこともあります。図1の様な時間軸の中で最初の頃, 即ち0年の近くにある種が, 例えば種1, 種2, 種3, 種4という種, それぞれ異なる種だと思って下さい。その0年から初期のほう, 例えば最初の10年とか, そのくらいの間に入ってくる植物っていうのをパイオニアあるいは先駆種と言っています。時間が経つにつれて, 最初の頃に生えていた種が次の種に変わって最後の種に変わっていく, その種の移り変わる過程を遷移と言います。最初に先駆種というのがあるわけですが, よく入ってくる種っていうのは, 噴火跡地などだと周りには植物っていうか, 供給起源が全くないわけですから, 遠くからやってこなくちゃならない。そのための種子を移動させる手段として, その遠くまで種を運ぶ能力のある風や動物に運ばれてくる |
ような種が最初の頃に入ってきやすい。次第にその中間の時期に入ってきまして, その頃を途中相とか中間期って言っています。最後には日本ではだいたい森林になるのですけれども極相と言われてる林になります。日本は, 高山帯とか非常に寒いとろを除けば, ほっとけばだいたい極相というのは森になるのですけれども, そういう意味もあってこちらっていうのは非常に日本中多くのところで調べられています。残念なことに噴火直後とか山火事直後にどういう植物がどのように入ってくるのか, それがどのように変化しているかっていうのはある意味で, 特に火山で研究例が非常に少ないっていうことで, この話を紹介させて頂きます。 極相の話っていうのは, 今日は全く出てこないので, 一応, 現在の遷移系列の中での今日のテーマであります多様性に関する事を一つだけ言っておきますと, 従来は極相が非常に多様な生態系であって貴重であると言われていましたが, 最近では中規模撹乱仮説が提唱され, 種数は, 遷移初期には非常に少ないのですが, それがだんだん時間の経過とともに増えていくと。その中間期っていうのは最も種間競争が激しい段階でいろんな種が, がんばって競争しあっていると, そういう状態の時は非常に種数が多い。ところが, その競争で敗れ去ったものが消えて形成される極相期は, 中間期に比べると, 多様性は低くなっている。だからある程度中規模のダメージを受けている, あるいは競争の激しい時期である, 遷移の中でも中間期っていうのが種数っていうのは最も多く, 多様であるっていうようなことがいわれています。遷移の中期から後期の話は全部やったことにしまして, 遷移の初期の話を続けたいと思います。 |
遷移は, どのように区分されているかと言いますと, 普通には1次遷移, 2次遷移という2つに分けられます。その違いは, 1次遷移は, そのある場所で植物が全くなくなってしまった頃から始まる遷移であり, それに対して2次遷移は例えば山火事跡とか, 耕作放棄地のような多少なりともその土地に植物が残っているところから始まる遷移のことです。2次遷移の始まりの例としては, この辺りでは知床にせっかく開拓に入ったのですけど, 結局その土地の生産力が低いためにみんな離
 農してしまって, もう見捨てられてしまった畑とか, そういうところですね。1次遷移はさらに2つ, 乾性遷移と湿性遷移に分けています。湿性遷移は沼とか湿原からの遷移で, 乾性遷移っていうのは乾いたほうの遷移で, 代表例として新しくできた島とか, 火山噴火とか, 溶岩が流れ出たその上での遷移のことを言います。今日まず扱うのは, ですから昔の教科書の言い方で言いますと1次遷移的なところの乾性遷移についての話です。
農してしまって, もう見捨てられてしまった畑とか, そういうところですね。1次遷移はさらに2つ, 乾性遷移と湿性遷移に分けています。湿性遷移は沼とか湿原からの遷移で, 乾性遷移っていうのは乾いたほうの遷移で, 代表例として新しくできた島とか, 火山噴火とか, 溶岩が流れ出たその上での遷移のことを言います。今日まず扱うのは, ですから昔の教科書の言い方で言いますと1次遷移的なところの乾性遷移についての話です。馬渡先生が高校教科書のことを話されていましたが, 図2は日本の高校3年生物の教科書が例えば10種類あったら10種類に載っている典型的な植生遷移, あるいは植物の遷移の基本的な概念図です。なぜこれを僕が今日最初に見せなくてはならないかというと, 高校の教科書ではよくこういうふうに紹介されているのですが, 有珠山ではこのような遷移をしていないのが一つの理由です。さらに, 外国で見てきた火山ではアメリカ合州国のセントヘレンズ山の山頂付近, ニュージーランドの北島火山群, さらにはピナツボというフィリピン, まさに |
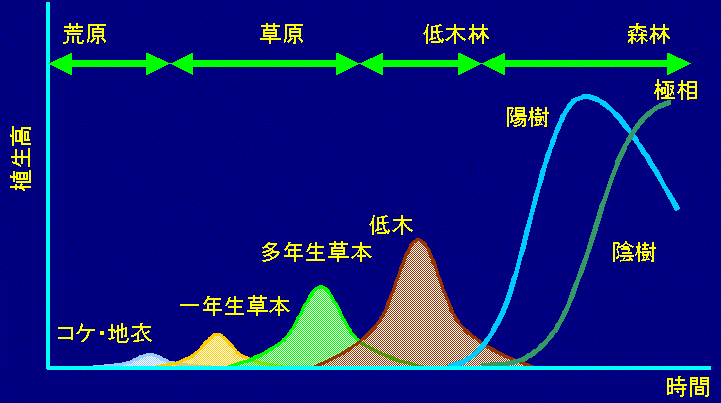 熱帯なのですが, それらの火山の噴火跡地などでも, この遷移系列の, 後ろの方っていうのはまだできていないのですけれども, 最初の方で既に教科書とは違うということを一つ言いたいのです。これが大学入試によく出るので, それに対して腹立っているのですけども。1次遷移と2次遷移っていうのは左端が時間の若い時で, 右に行くにつれ時間がどんどん経過していると考えて下さい。その遷移の最初に何が入ってくるかっていうと, 1次遷移では苔, 地衣, 藍藻, こういったものが入ってくると説明されています。2次遷移では最初に1年生生草本,その生活史の非常に短い,春出たら秋には花をつけてしまうような非常に短い生活史をもっている植物が最初に入ってくる。その理由を言うと, 火山の噴火跡とか溶岩上では非常に栄養分が乏しく種子植物, 1年生草本, 多年生草本, 低木, 陽樹,陰樹っていうふうに並べてますけども, このような種子植物は貧栄養土壌の上では非常に成長が困難である。だからそういう連中は, 溶岩上とかには最初に入っていけないので, まずほとんど土壌窒素がなくても生活できる苔, 地衣, 藍藻などのようなものが地表面に定着するというふうに説明されています。それらがある程度土壌を形成すると, やっと1年生草本植物が侵入でき, それから時間が経過するにつれて極相である陰樹の林になるというふうに説明されています。山火事跡とか耕作放棄地の遷移は2次遷移って言いうわけですが, そちらのほうっていうのは, ある程度土壌ができているのでコケなどが優占するステージは必要ではなく1年生草本から遷移が始まるというふうに説明されているわけです。しかし, 有珠山ではそうではありません。
熱帯なのですが, それらの火山の噴火跡地などでも, この遷移系列の, 後ろの方っていうのはまだできていないのですけれども, 最初の方で既に教科書とは違うということを一つ言いたいのです。これが大学入試によく出るので, それに対して腹立っているのですけども。1次遷移と2次遷移っていうのは左端が時間の若い時で, 右に行くにつれ時間がどんどん経過していると考えて下さい。その遷移の最初に何が入ってくるかっていうと, 1次遷移では苔, 地衣, 藍藻, こういったものが入ってくると説明されています。2次遷移では最初に1年生生草本,その生活史の非常に短い,春出たら秋には花をつけてしまうような非常に短い生活史をもっている植物が最初に入ってくる。その理由を言うと, 火山の噴火跡とか溶岩上では非常に栄養分が乏しく種子植物, 1年生草本, 多年生草本, 低木, 陽樹,陰樹っていうふうに並べてますけども, このような種子植物は貧栄養土壌の上では非常に成長が困難である。だからそういう連中は, 溶岩上とかには最初に入っていけないので, まずほとんど土壌窒素がなくても生活できる苔, 地衣, 藍藻などのようなものが地表面に定着するというふうに説明されています。それらがある程度土壌を形成すると, やっと1年生草本植物が侵入でき, それから時間が経過するにつれて極相である陰樹の林になるというふうに説明されています。山火事跡とか耕作放棄地の遷移は2次遷移って言いうわけですが, そちらのほうっていうのは, ある程度土壌ができているのでコケなどが優占するステージは必要ではなく1年生草本から遷移が始まるというふうに説明されているわけです。しかし, 有珠山ではそうではありません。
|
|
ここで, なぜ火山遷移の研究は重要なのかという意味づけをしておきたいと思います。世界における火山の分布及び地震の分布を見ますと, 日本は環太平洋火山帯の中のセンターというか, 非常に火山の多い, 集中しているところである。今世紀大噴火を行った火山の多くがこの火山帯に属していまして, 先程言いましたセントヘレンズ山というのはアメリカ西海岸にあり, ピナツボ山はフィリピンにあり, またニュージーランドの北島というのも非常に火山の多い所です。日本の中ではどうかと言うと, 北海道にはA級火山と言われています, 24時間常時監視体制が必要である極めて危険な火山が, 雌阿寒, 十勝岳, 樽前, 有珠山, 駒ヶ岳と5つあります。記憶に新しいのでは去年, おと年と駒ヶ岳が小噴火をしました。今日紹介します有珠山は1977年~78年にかけて噴火した火山です。十勝岳の噴火は皆さんには記憶に新しいものと思います。北海道に |
おける火山噴火の特徴の一つは, 殆どの火山の噴火時には火山灰, 軽石の噴出を伴います。場合によっては当然サージとか, 泥流, 当然火山灰, 軽石, 泥流が出るのですけども, そえらによって大災害がもたらされやすいっていうのが一つの特徴です。さらに, これらの噴火によって北海道全域に火山灰, 軽石を降らせていますので, 例えば, これは釧路湿原などで穴を掘ってみるとよくわかるのですけれども, ああいう所でも必ずそのある深さのところに火山灰層が出てくる。というように, 北海道の植生っていうのは, 一見すると非常に成熟した林であっても, 多かれ少なかれその火山灰とか軽石という火山噴火の影響を受けて成立しているっていうのが特徴になっているわけです。したがって, 火山噴火の植生に与える影響を初期から調べることは遷移機構を知る上で非常に意味のあることと考えています。 |
|
有珠山の噴火の歴史ですが, 地球物理学的には1977-1982年にかけて有珠山は火山活動が認められています。始まりは1977年のこの軽石の大噴出噴火, 7, 8月の7日から14日にかけて, ここから噴火が始まるのですが, 地球物理学的には82年まで火山性微動とかそういうのが続いていて, 火山活動はありました。ただし我々植物をやっている人間にとりましては, 植物が影響を受けるっていうことが火山性微動からくるっていうことはまず考えられませんので, 植物に被害を与える火山噴出物が出るのをやめた時, そこまでを噴火の期間としています。それはいつかと言いますと, 1978年10月27日に小噴火があって火山灰を噴出したっていう, その段階をもって一応火山噴火の終りとします。というわけで, 火山噴火の生態系に対する影響は77年-78年にかけてのおおむね1年半から2年間であるということになります。その78年っていうのを遷移の始まり, 開始0年ということにして考えてよいと思います。 よく噴火当初はどのようだったかと聞かれるのですが, 残念なことに, その時( 77-78年)は, 出身の茨城県の実家で高校3年生でしたので自分では見ていません。むしろ札幌に来てみんなに, 札幌でも車の上に火山灰が積もっていたとか, そういう話を聞くたびに, ああ, そういうものなのかなあ, という程度で自分でこうだったと言えないのが残念です。しかし, 噴火直後の航空写真を見ると当時の状態が非常によくわかります。図3にありますように, 第4火口という4回目の大噴火をしたところですが, それが有珠山における1年半の噴火の中で最大だったのですが, その時にできた火口を示しておきます。直径200 m位だと思うのですが。また, 山頂部から外輪山にかけてガリーという, 沢, 日本語で言うと沢としか訳せないと思うのですが。大きな雨とか雪解けの水で削れてできた沢が形成されています。火口源内には大小のガリーが, あちこちにできていました。ガリーができているってことはこの辺には非常に火山灰が厚く堆積していたということです。だいたい火口源辺は火山灰で植物がほとんどなくなってしまった。ところが, 外輪山辺は噴火で死ななかった木が生き残っていたわけです。有珠山は外側を外輪に囲まれまして, その中に二つの山頂をもっています。一つが大有珠と言われている所で, もう一つが小有珠と言われております。噴火前はこの山頂部も, 緑々していたのですが, それが第四火口, それ以外の部分でも何箇所か, 10回くらいで噴火しているのですが, その時に火山灰が積もっったり, 直後に雨が降るとその雨の重みで枝が折れたりとかして, 木が死んでしまったり, あるいは爆風で植物が死んで殆ど裸地状態になってしまったと, ここから植生回復が始まったわけです。先程言いましたように, 噴火直後, 僕はまだ高校生から大学生にかけてのときで, その頃のデーターは自分では取っていませんので噴火直後のデーターは, 帯広畜産大学の紺野先生のデーターによるものですが, 火口原から外輪山にかけて確認された植物は, 次の4種でした。この報告書では残念なことに調査区の大きさ書いてないので, どのくらいの面積でこうだったのかがよくわからないのですけども。ともかく生えていた植物ってのは若干なりとも植物の生きていた外輪山でもオオブキ, エゾノギシギシ, スギナ, アザミの仲間の4種が生えていた。6つの調査区を作ってその中に4種があったにすぎない。噴火の時にギリギリ爆風の被害を逃れた植物が, 多少なりともある辺りが外輪山って言っている部分です。火口源と言われる平坦部に火山灰が数メーター堆積しまして植物が殆ど死んでしまった辺りです。火口源に至っては, 2調査区しか調べてないのですけれども, そこでは非常にまれにしか出てなかったという意味でエゾノギシギシだけがちょこっと, だから多分実生, 芽生えがちょこちょこっと生えてだけっていうような状態だったっというように考えられます。だからほとんど火口源っていうのは全く何も生えていない状態から植生回復が始まっているといえます。 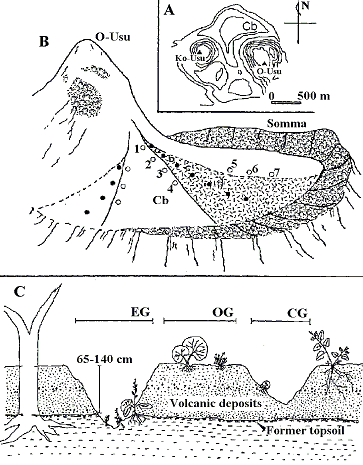 さらに今のが噴火直後, 79年に調査された, すなわち噴火から1年経過した段階なのですが, 今度はそれから噴火3年後ですか。3年後に航空写真と実際現地を歩いた調査結果をもとに, フランスからの留学生のアン=リビイルという方が作った植生図をもとに見てみます。第四火口がありまして, その周辺のいっぱい植物が生えているところが外輪山です。火口源では, ここは殆どまだ, 若干の植物の侵入は認められるけども, まだ定着には至っていない。おそらく殆ど芽生えばっかりでそれが雨とかで, 雪とかで流されて死んじゃってることの繰り返されていった状態だったものと思われます。それに対しまして外輪山では非常に植物がたくさんびっちりと生えているっていうふうに, そのやはり噴火の時に絶滅してしまった場所と, 多少なりとも生き残っていた場所とでは植生回復の速度が全く違うというのがわかります。
さらに今のが噴火直後, 79年に調査された, すなわち噴火から1年経過した段階なのですが, 今度はそれから噴火3年後ですか。3年後に航空写真と実際現地を歩いた調査結果をもとに, フランスからの留学生のアン=リビイルという方が作った植生図をもとに見てみます。第四火口がありまして, その周辺のいっぱい植物が生えているところが外輪山です。火口源では, ここは殆どまだ, 若干の植物の侵入は認められるけども, まだ定着には至っていない。おそらく殆ど芽生えばっかりでそれが雨とかで, 雪とかで流されて死んじゃってることの繰り返されていった状態だったものと思われます。それに対しまして外輪山では非常に植物がたくさんびっちりと生えているっていうふうに, そのやはり噴火の時に絶滅してしまった場所と, 多少なりとも生き残っていた場所とでは植生回復の速度が全く違うというのがわかります。それで1983年, 噴火5年後から, 私は有珠山の植生回復を調べようということで調査を始めたわけです。さっきの噴火3年後にアン・リビイルが植生図を作成したときと殆ど同じ状態で, 噴火6年後ではこの外輪山というのは殆ど植物でびっしり覆われていた。これは主としてオオブキとか, オオイタドリっていう大型の多年生草本です。それに対して火口源っていうのは火山灰に覆われ火山灰の上には殆ど何も生えていないという状態でした。 1983年に山頂部から外輪山にかけて2 m × 5 mの調査区を適当に設置して, 今まで調査を行っています。そこで, どのように植物が変わっていくのか, どういう植物が侵入するのかという話しをしたいと思います。まず, 有珠山火口原の地形区分を説明しておきます。有珠山では火山灰は何 mも積もり, 雨とか雪で火山灰が削れてガリーができます(図3)。そのガリーのでき方から大きく生息地あるいは地形的環境は3タイプに分けられます。1つは, 火山灰の下にある噴火前の土にまで土壌侵食が進みガリーが大きくなって削れ噴火前の土まで到達し, 噴火前の土が露出しているガリーがあります。それに対し, まだ旧表土, 噴火前の土までガリーが達してないところがあります。また, ガリー侵食の殆どない平坦な部分の, ガリーの外側, の3つのタイプです。 遷移初期段階の植生を決定づけるのに重要なことの1つは, どこからどのような植物がくるのかという問題があります。火山噴火後は, 殆ど植物が無いので, 周りからやって来るしかない。どのような植物の種子がくるのかを調べるには, 種子を捕まえればいいということで, よく使われるのがシードトラップと言う方法です。ある大きさの枠の中に, 網をはり, 種子を網の中に引っかかるようにしておきます。それで, この網を1週間とか2週間ごとに札幌と洞爺を行ったり来たりして, 交換して実験室でこの網の中にゴミもたくさん入っているのですが, その中から種子を取り出し種を同定するわけです。 まず, 先程, 門田先生が言っていました種子の散布型について説明したいのですが, オオブキやオオイタドリ, そういう種っていうのは風散布と言われ, 風により種を飛ばし移動する仲間です。それに対し, 子供の頃に友達同士でくっつけあって遊んだ種子がいっぱいあると思うのですけれども, ああいうのは動物散布と言われているやつ。それに対して自分ではじけて種を飛ばしたりするやつを自発散布と言っておりまして, あとヤシの実みたいに水に乗って動くやつっていうのは水散布って言われています。種子以外に植物体を移動させる手段として栄養繁殖といのもありますが, 大きく種子の散布系っていうのは, この4つに分けられています。それに対し種子を移動させるのに特別な器官をもたないものは, 昔は重力散布と言われていました。しかし, 最近ではドングリなどは, どう分布域を変えるのかという話になった時に, 重力散布ではとても移動に貢献しているとは思えない。そこで, 最近はネズミが種子を冬越しのために土の中などに貯めておき, 食べるのを忘れた種子が発芽する。そして, 分布域の変 |
化に貢献しているという説があり, 厳密な意味での重力散布は, 考える必要がないという話もでてきていますので, 重力散布については, ちょっと保留しておきます。それ以外はこういう形で種子の分散型を分けている, 生態学上は分けていると考えておいて下さい。実際に風散布の種子は, こういうのが網の中に引っかかるわけですね。ひとつのタイプとしては, ヤナギとかポプラの仲間やタンポポみたいに綿毛でポワポワと空を飛んでいるやつ。もう一つはカエデの仲間で, カエデなどは綿毛のようなものをつけ種子を飛ばすのではなく, 羽をつけ, この羽で飛ぶという, こういうタイプが風散布の種です。 有珠では, 1984年と1985年に外輪山と火口源にシードトラップを置き, どの位の種子がくるのかを調べました。すると, やはり植生回復が早かった噴火の被害をまぬがれ多少なりとも植物が生き残り, それらがあっという間に大きくなった外輪山で非常に沢山の種子が取れました。オオブキとかオオイタドリ, あとエゾノコンギクといった風散布性のものがたくさん取れた。さらに火口源では数が少ないのですが, 同じような仲間の種子が多少なりとも飛んできくるということもわかりました。それ以外にヤナギの仲間とかドロノキ, イネ科の仲間, そういう植物の種子もトラップにかかりました。こういう植物は外輪山より外側から風で入ってきていると考えられます。 しかし, それだけでは有珠山に生えている植物の全種数からいうと非常に少ない。特にどう考えても風で数キロも種を飛ばすとは思えない植物が有珠の火口源ではけっこう優占している場所があり, どこから来てるのかを考えてみると, 1つはやはり噴火前の土があやしいっていうことで穴を掘ってみたわけですね。2 mも掘ると噴火前の土にとどきました。オオブキの場合には, 火山灰が50から60 cmの深さであれば十分地下器官を繁殖させ, 地表面まであがって来られます。色々掘ってみたら何種かは, 噴火前の土があると, そこに地下器官の塊があって, それがずっと伸びてって地表面でもう一回その塊を作って, そこから葉っぱをたくさん出していました。特にオオイタドリなどは1.5 mくらい火山灰の厚さがあっても, その噴火前の土のところに生き残っていたその塊から, ちゃんと地表面まで上がってきて大きくなれるようです。他の多年生植物でもたとえ長距離, 種子を移動できないやつでも浅いところをうまくみつけて, そういうところから十分栄養繁殖で回復していることがわかりました。 もう1つ調べていて不思議だったのが, 旧表土の出たガリーと旧表土の出てないガリーに分け, その2つの間に出現している植物種を比較しますと, 旧表土の出たところでだけたくさん出てくる種が10種以上ある。こいつらは, なぜ旧表土のあるところに完全に依存して定着しているのかという疑問がわきます。1つ考えられているのは, 旧表土の中で種が生きていたのではないかっていうことで, 実際に昔の土を掘って取ってきまして, 実験室でその土に水をやり育てますと, 5種しか発芽しませんでした。これでは少ないので, 10何種を全部土の中かから取り出そうということで, 全部とるのにどんな方法があるか色々と調べていた, 僕が博士課程にいた時の教授の吉田先生から, 土を50%炭酸カリウムという水溶液に混ぜ, 遠心をかけてやると種子が浮いてくるのて, それを濾しとり, 同定すればいいということで, この方法でやると実際たくさんとれました。土を, 旧表土噴火前の土をとってきたのが1987年ですので, 噴火後9年目になります。その時に深さが60 cmから1.4 mくらいのところにある噴火前の土をとってきて, その中の種子で生きているものをピックアップしていくと, 少なくとも25種が生きていました。そのうち少なくとも12種は実際に旧表土の出たガリーの中だけで特異的に定着している種で, それらがこれだけたくさんあったということです。例としてはエゾノギシギシ, ヒメスイバ, イヌタデ, ミチヤナギ, スズメノカタビラ, スズメノテッポウ, こういう植物でした。これらは明らかに噴火10年を経過しても, 噴火前の土の中に生きていることのできる種から供給されているということがわかりました。この続きとして去年が噴火20周年なので, うちの学生に20年後の土とってきて, どのくらい生きているか調べてみてくれって言ったら, 20年後の土でも, まだたくさんの種が生きているっていうことで保存状態は非常にいいです。チャンスがあれば, これらの種っていうのが地表面に出てきて植生回復, あるいは遷移に寄与することは十分に有りうると思います。 実際にそのいろいろな供給源からきている植物が1つのパッチを作っているのを見ますと, 栄養繁殖起源のオオブキ, 埋土種子起源のシロツメクサ, 埋土種子起源のエゾノギシギシ。有珠山では砂防のためにヘリコプターから種子をまいていて, そのような人工播種からきているオオウシノケグサ, それとシードトラップで捕獲されたヤナギの仲間などの木本植物といういろいろな植物供給起源から来た植物が, 多様な群落を作っています。 まとめてみますと, 有珠山では植物供給起源は栄養繁殖, オオブキとかオオイタドリといった大きな活力の高い植物, 長距離種を運べる移入からきてるもの, 航空実播, そのヘリコプターでまかれた種からくる人工播種, それと埋土種子から, という4つの植物供給起源から植生回復が始まっています(← 表1)。 最初の方で今の高校の教科書では1年生草本から植生回復が始まるというふうに, っていうか最初の頃には一年生草本の優占するステージっていうのがあると言われているといいました。これは一年生草本, 多年生草本, 木本植物が, ガリーの外部, 旧表土の出たガリーの内部, 旧表土の出てないガリーの内部で何種類あったかをまとめてるのが表1です。さらに, それらが1年生植物の供給起源が栄養繁殖, 移入, 人工播種, 埋土種子から何種きているかをまとめた表なのですが。1年生植物っていうのはその殆どが, 埋土種子, すなわち旧表土中にあった種子から供給されています。ほかの植物供給起源, 例えば移入, 外輪山の外側から風とかに運ばれてきた種はわずか1種しかありません。このように, もし旧表土から埋土種子が供給されてなければ, 一年生植物はわずか1種しかない遷移の初期段階となります。そういう意味では, 埋土種子という植物供給起源がなければ一年生草本期っていうのは全く形成しようがないっていうことが言えます。その意味で植物の供給起源が遷移の初期段階に何が出現するのかを大きく左右しています。 このようにして, どのような種がどこに入って来るのかっていうのは供給起源によってかなり決定されているということが言えると思います。次の段階として, では入ってきた植物っていうのは, どう大きくなってきていけるのか, あるいは逆に消えてしまうのか, というのを全体でみてみます。僕が調査を始めた83年から97年, 一昨年までの年次変動を説明します。まず種数の年次変動ですが。ハビタット別にみますと旧表土の出たガリーで種数が毎年ずっと多くて, ガリーの外側, 旧表土の出てないガリーっていうのはだいたい10種くらいでもって97年まで種数が停滞しています。この種数の差っていうのは主として埋土種子から供給された植物があるか, ないかによるものです。最近ではだんだん収斂してきてるっていうのがわかります。全体として種数はこのくらいですっと推移しています。このことからも, 植物供給起源にどの位種が含まれているかが重要であることがわかります。 植物の大きくなり方ですけども, 調査を始めた噴火5年目である83年から97年までの, 各年の値を見てみます。ここでいう被度とは, 上から見た時にあるところ, 例えば調査区, を植物が何%覆っているかってという意味です。例えば, 被度オオブキの50%となっていれば, オオブキがその調査区面積の半分を占めているという意味になります。その各種の被度の合計値をまず見てみます。おおざっぱにいえば, 噴火10数年を経過した1990年, 91年あたりで100%というふうになってるっていうのは, だいたい上から見た時にそのプロットが, 植物でおおむね覆われている。100%を超えてるっていうことは植物の層っていうのが2層以上できている場所となっているということです。そのくらい非常に順調に被度は増え続けています。 次に, どの種の被度がすごくよく大きくなっているのかというと, まずオオイタドリという, タデ科の超大型, 僕の身長よりも大きくなるという植物なのですけども, その年次変動を見てみます。噴火終了初期には外輪山とかでもすごい優占してるように非常に順調に大きくなってるんですけども, 89年くらいから頭打ちになりまして, もうそれ以上増えてません。定着地としては旧表土の出たガリーでは全く定着できないで, それ以外のとこで優占しているっていう特徴があります。もう1つやっぱり地下部を掘ると, 比較的深いところから植物が栄養繁殖で回復できる種であるオオブキも着々と被度を増やしている。オオブキの場合は, オオイタドリはこの辺で頭打ちになっていますけど, 今のところちゃくちゃくと面積を拡大していまして, 特にガリーの内部で優占してるっていう特徴があります。その結果として, 大型多年生草本が, 今現在優占しているステージになっているわけです。ところで, 有珠山の場合には苔っていうのが, 殆ど全く生えていませんでして, さらに一年生草本というのも旧表土の出ているところ以外では殆ど出てきませんでした。最初からオオブキとか, オオイタドリっていう多年生草本から植生回復が始まるっていうのが特徴になっています。 |
|
次にいつになったら森や林になるかの目安である木本植物の増加ですけが, これはドロノキというポプラの仲間である植物の被度の年次変動を見てみます。その被度の増加は, 多年生植物, オオイタドリとかオオブキに比べますとゆっくりなのですが, 着実に増えています。多年生草本のオオイタドリはだいたい合計の被度が30%くらいで頭打ちになっていますが, 木本植物のドロノキの方は着々と被度を拡大しています。また, 木の高さも数 mに達しており, もっとも大きな草本植物であるオオイタドリの背丈を追い越しつつあります。極めておおざっぱな予測にはなりますが, あと10何年くらいでドロノキのほうがオオブキよりも被度では多い状態になるでしょう。したがって, あと10何年かくらいでもって, 森的な景観になるっていうように非常に早い植生回復をしている火山であるということが言えると思います。 旧表土の出たガリーの中でも, やはり植物にびっしりと覆われている, 非常にいい状態となっています。噴火直後っていうのは, 調査を始めた頃ってのは本当に最初にクイを40本くらい調査用に打つと, 次に調査に行った時には10本くらい流されてるっていうくらい, 雨や融雪水による土壌移動が激しかったのですが, 徐々に安定してきました。また, 土壌も目で見た感じでは少しずつ形成されつつあります。 今の有珠山全体がどういうような植生になっているのかを, 調査区の中で調査期間に生えていた全種のデーターを基にDCAという序列化手法を使って分析 |
しますと以下のような結果が得られました。ここではXY軸を用いた2次元グラフ上で視覚的に説明したいと思います。まず, アカザ, セイヨウタンポポ, エゾノギシギシ, イヌタデなどは全て埋土種子起源の植物ですが, これらの種がたくさん出てくる調査区は, X軸上で高い値をY軸上で低い値を示しています。それに対しアマニウ, アカソ, シラネワラビ, エゾヤマザクラ, シラカンバのような木本生植物, あるいは森林性の植物はX軸上で低い値をY軸上で高い値を示しています。そして, これらの種がたくさん出てくる調査区はやはりX軸上で低い値をY軸上で高い値を示しています。これらの種の得点と各調査区の15年間の動きをみますと, X軸に沿い変動している調査区は埋土種子集団起源の種の出現に引っ張られているグループといえます。それに対しY軸に沿い動いている調査区は, 森林性の種の増減に左右されているもので, どちらかと言うと遷移が進んでいる, より森林的な植生に近づいているものと言えます。それに対して得点がX軸Y軸ともにほとんど動かない調査区は今でもその植生の変化が小さく遅いグループと言えます。まとめてみますと今のところ調査した範囲の中では埋土種子集団によって植生が規定されている。しかし, 森林にはあまり進んでいかないような調査区, それと植生の変化が停滞してしまったグループ, および除々に遷移が進行し森林化に向かっているグループ, という3つの植生の型があの狭い火口源の中で分化を始めているという状態にあると考えられます。以上です。 |
(北海道大学大学院地球環境科学研究院・温暖化影響評価部門)
地球温暖化の原因が人為かどうかはさておき、近年、地球規模で気温上昇が起こっていることは否定できない。殊に、温暖化は、極地生態系において大きな脅威となっている。すなわち、永久凍土層の存在が、極地生態系の構造を規定する主要因であるが、その消失が懸念される。アラスカ、フェアバンクス近郊では、永久凍土を欠如する南側斜面ではシロトウヒ林が、永久凍土が存在する北側斜面ではクロトウヒ林が発達する。さらに、アラスカでは、森林火災がこれまでにない大規模なものとなっている(これも温暖化が原因とされる)。そこで、アラスカ内陸部で2004年に発生した大規模火災地内に永久調査区を設け、追跡調査を開始した。ここでは、少なくとも遷移初期にはクロトウヒは、更新できていないこと、そして回復にはミズゴケの定着と凍土の発達が重要であることを指摘したい。