(2026年6月27日更新) [ 日本語 | English ]
HOME > 講義・実習・演習一覧 / 研究概要 > 小辞典 > 遷移
遷移概念 (concept of succession)1863 Kerner: 初めて極相概念(climax concept)紹介1899 Cowles: 遷移(succession)に関する(恐らく最初の)研究 1904 Clements: 遷移理論(succession theory)提唱 1899 Cowles, 1916 Clements
遷移(植生連続) (ecological) succession = 植物群集の時間的推移: 植物群集が時間と共に種の交代によって(自然に)変遷する過程。遷移といえば普通は生態遷移のこと 1916 Clements: 発生(出生) birth - 成長 - 成熟 - 死 death = cyclical change 1923 Nichols GE: 地文的極相 topographic climax
地形・立地条件 topography に支配される植生安定状態 (≠ 土壌的極相) 時間の経過に伴う群集の変化 図. 遷移の概念図 (Watt 1947) 輪廻説 cyclic theory再生複合体 regeneration complex: 極相に向かう様々な遷移段階の群集からなる自律的遷移・他律的遷移 autogenic and allogenic succession自律的遷移(自発的遷移): 植物の環境形成作用により起こる遷移
分解的遷移 degradative succession: 有機物分解過程での群集変化
(セルロース・リグニン)完全分解して遷移終了 生物的極相(他極相説): 生物要因による極相 = 準安定相(単極相説)
⊃ 人為極相、火成極相 |
[ 火山遷移 | 有珠 | 駒ヶ岳 | クロノシークエンス ] 用語 [ 亜極相, 環境傾度分析, 気候的極相, 極相, 極相種, 極相群集, 極相パターン説, 極相林, 後極相, 湿性遷移, 前極相, 土壌的極相 ] 系列 sereEx. 砂丘: 不安定(植物無) → 半安定(植物少数) → 固定砂丘・安定(植物数増大)
一般的植生遷移モデル general succession models 先駆種 (pioneer): 主に遷移の初期に現れる種 極相種 (Bazzaz 1979) 遷移に関する生理生態的特性Table. Physiological ecology of plant succession
属性: 遷移初期植物 ⇔ 遷移後期植物
先駆種的 極相種的
種子 散布期間: 長い ⇔ 短い
二次休眠: 普通 ⇔ 普通ではない?
種子発芽 促進 光: ○ ⇔ ×
温度変動: ○ ⇔ ×
高NO3-濃度: ○ ⇔ ×
種子発芽 抑制 赤外光: ○ ⇔ ×
高NO3-濃度: ○ ⇔ ×
樹種 陽樹 ⇔ 陰樹
光飽和点: 高 ⇔ 低
光補償点: 高 ⇔ 低
低照度有効性: 低 ⇔ 高
光合成速度: 速 ⇔ 遅
呼吸速度: 速 ⇔ 遅
蒸散速度: 速 ⇔ 遅
気孔及び葉肉抵抗: 小 ⇔ 大
水輸送抵抗: 小 ⇔ 大
訓化能力: 大 ⇔ 小
資源制限からの回復: 速 ⇔ 遅
過激環境縮小能力: 大 ⇔ 小?
生理学的反応幅: 広 ⇔ 狭
資源取得速度: 速 ⇔ 遅
物質分配可塑性: 大 ⇔ 小
|
|
一次遷移 primary succession: 土壌中を含む全群集が消失した状態からの遷移 (植物起源: 移入)
湿性遷移 mesic succession: 湖の陸化過程や中州など水の量と動きに決められる遷移 火山噴火・新島 → 火山遷移 (volcanic succession) (露崎 1993, 春木・露崎 2008)
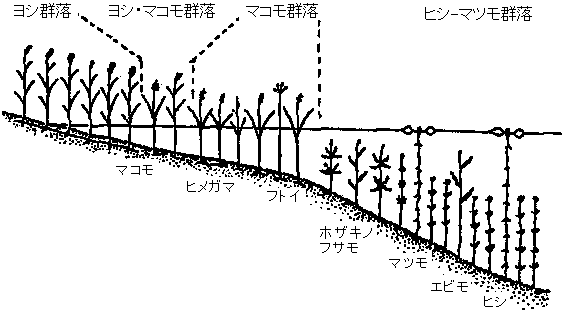
塩性(乾性)遷移 halosere: 干拓地、塩田跡、干潟等の高浸透圧海水に洗われる基質で始まる → 陰樹林完成すると安定 = 安定期(極相climax) 陰樹: 日照量少い環境でも生育に耐える樹木。Ex. 針葉樹 = ヒバ、モミ、トウヒ、ツガ類、広葉樹 = ブナ、シイ、カシ類等 ⇔ 陽樹 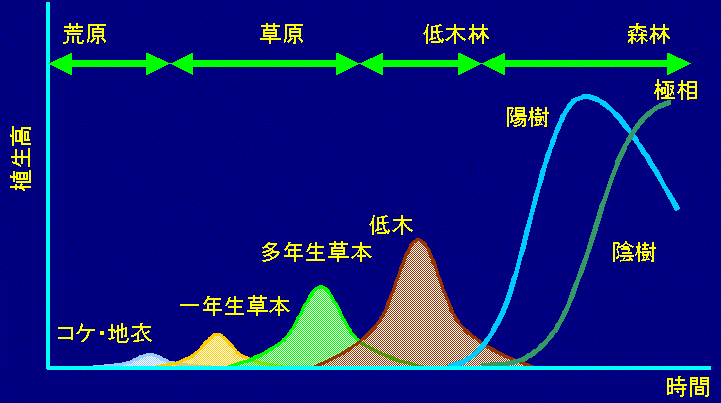
図. 乾性一次遷移の典型的とされる模式(Whittaker 1975)。時間の経過につれ植生高が高くなる |
表. 乾性遷移系列 xerarch。*: 温帯 - 亜寒帯
群集 草原 低木草原 陽樹林 混交林 陰樹林
年 4-5 5-20 25-100 > 150年 亜寒帯 サンリンソウ シラカンバ オオシラビソ イワイチョウ トドマツ 北海道* スゲ シラカンバ 広葉樹 エゾマツ ブナ トドマツ ヤチダモ ハンノキ 暖帯 ブタクサ ススキ アカマツ シイ メヒシバ イタドリ クロマツ カシ 一年生 多年生 コナラ 二次遷移 secondary succession: 群集が除去された状態からの遷移 (移入*・栄養繁殖・埋土種子)
スキー場放棄・森林火災・泥炭採掘・石炭採掘 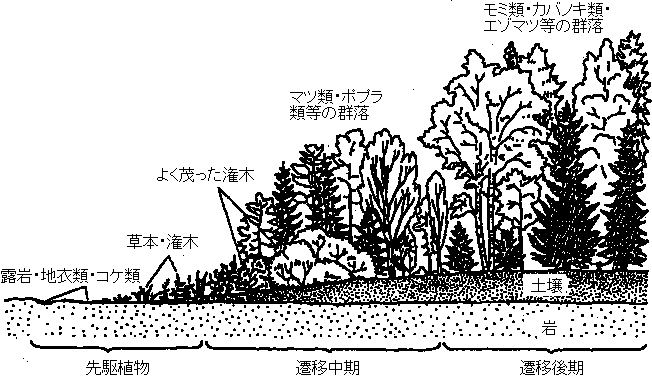 図4.6 生態遷移の例. 場所により図中のいずれかの先駆植物から遷移が始まる(松本 1993) |
|
基質中に植物種子・胞子・根系等の繁殖体を含まず、土壌形成もない
[ 昔言われてた遷移系列: 一年生草本のステージはないのに注意 ] 小規模 small scale
|
大規模 large scale
|
湿性遷移 (mesic or hydrarch succession)群集遷移の一形態で、遷移の開始期に湿った環境から始まる遷移の総称である。その遷移は、まず沼沢地化といわれる湖沼の形成があり、ついでそれらの沼沢地が、周囲からの土砂の流入、植物の侵入とその遺骸の堆積、により陸地化していく2つの過程でなされる。従って、植物相の変化は、植物プランクトンから水生植物に変化する過程を経由し、ついで陸生植物が表れるという流れとなる。なお、この遷移系列を湿性遷移系列と呼ぶこともある。環境傾度上では、(地下)水位傾度が時間の経過とともに高いところから低いところに向かう群集変化と見ることができる。陸地化後は、乾性遷移の途中相のいずれかの部分に組み込まれる。ただし、ミズゴケ湿原や湿性ハンノキ低木林などは、水位が安定し、そこに長い年月にわたり高木の侵入がなければ、土壌的極相とみなせる。(露崎 2010)湖沼(湖岸、中州、河岸)、池など水分の多い基質の陸地化により進む → 湿性遷移系列 hydrach 0. 沼沢化 paludification, swamping: 乾地や森林から湿原形成されること 1. 湖沼
湖 lake: 最深部に植物生育していない → 沼沢地: 最深部まで挺水植物生存する
貧栄養湖 - 土砂堆積等で浅化 - プランクトン等↑ - 透明度↓ - 富栄養湖 プランクトン、水生植物由来有機物からなる(青白色の)湖底堆積物 栄養物と酸素多く有機質栄養に富むか、過剰な水にできる
藻類骸泥 algal gyttja 腐植泥 dy: 水中で形成された泥状酸性腐植土層 → 基層の上に堆積
生物学的に極めて不活性で、腐植ゲルの無定形沈殿物を多量に含む褐色の水底で生成される |
表. 富栄養湖と貧栄養湖
特徴
富栄養湖
貧栄養湖
堆積が更に進み湿地形成
さらに草原となり、やがて低木、陽樹が入り込んでくる 5. 陽樹林 (以降は乾性遷移と同じ系列となる)6. 混合林 7. 陰樹林(極相) |
|
林野火災、森林伐採、田畑休耕、建設目的で裸地化した所等、既に土壌がある程度準備され、土壌の中に種子(埋土種子(露崎 2008)、胞子、根系などが用意されている場所である。人工植生や造林地は二次遷移から脇道にそれ、任意の樹種を裁植することであるが、自然の遷移の進行速度に比べ早く植被を得ることができる。人工的に植林を含める(s.l.) 二次遷移系列 secondary successional sere: 二次遷移に見られる系列 二次植生 secondary vegetation: 撹乱後に再生した植生 (s.l.)
≈ 二次遷移系列に属する各群落 Ex. シラカンバ林(ダケカンバ林) - 撹乱 (Ex. 森林火災後一斉更新) Ex. 薪炭林: コナラ、クヌギ、ミズナラ - 萌芽林
カシ林・シイ林も同様に薪炭林(二次林)であることある
一斉林となること多 + ササ林床発達
アカマツ林: 屋久島 - 青森県 (瀬戸内海沿に多) 日本温帯域 二次遷移日本西南部 森林伐採跡地先駆期: 一年生草木群落 (ベニバナボロギク): 1-2年ススキ群落 陰樹林: シラカシ,シイ,タブなどからなる萌芽林 = 極相 普通には雑木林coppiceと言われる → 陽樹林を欠く 日本西南部山地に見られる雑木林はこのタイプの森林が多い。繰り返し伐採し、天然更新を行なっているとクズ(Pueraria lobata)などの蔓植物の侵入が著しくなり、植林をした際に蔓植物の除去が大きな作業の一つになる。
田畑休耕地 abandoned pasutreブタクサ, エノコログサ等: 一年生植物ヒメムカシヨモギ等Erigeron: 越年生植物 セイダカアワダチソウやススキ等: 多年生草本(同時にヌルデ、クロマツ、ナワシロイチゴ、等木本植物も侵入) 陽樹林期 二次遷移は既に土壌が完備され(Ex. 田畑跡等は高窒素分)一次遷移より進行速い。森林伐採や田畑放棄直後に一年生草本優占群落が成立するのは、以前から植生が存在した段階で土中に埋土種子集団として貯蔵されていたから |
二次遷移初期にドングリ(ブナ科種子で、森林優占種を占めるブナ、シイ、アカガシ、ウラジロガシ等を含む)を植えると育つが、天然には遷移初期に出現しない。これは種子散布様式に糸口がある Def. マント群落 mantle community: 林縁内側にできる外套状の群落
道路開設等の人為干渉で形成された二次的植生
日当たり良 → 陽生草本・低木優占 種子生産 seed production先駆種としての種子の条件 → r-淘汰的
全てアロメトリー成長関係 allometric relation となる
ハルタデ: A = 2.843, B = 4.061
Ex. ヒメジョオン優占区: 66個体/(50 × 50 cm²) → 約22万粒の種子生産 |
|
極相: 自然は変動する - 変化: 唯一の真実/事実 (Bergson「生命とは何か」)
環境問題にこの事実を考慮しないため誤った自然保護が行われがち 遷移系列上で、通常は最後に位置する群集を極相という(☛ 遷移)。ただし、気候的極相説(単極相説)の立場と、土壌的極相説(多極相説)の立場では、その極相観は異なるが、多極相説の方が、より多くの現象をよく説明でき、さらに極相パターン説の出現にともない、気候的極相概念を用いることは廃れている。極相パターン説からは、明瞭な極相を定義する必要はなく極相は時間とともに変化する環境経度に沿って連続的に変化した最終段階と考える。いずれにしても、極相は、相対的には遷移初期から中期に見られる撹乱やストレスの高い群集に比べて、種の構成や群集構造の変化が小さく、平衡状態に達したか、それに近い状態にあると考えられる。実際には、極相と認識できる群集は、ギャップ形成等の撹乱により部分的に種の入れ替わりが起こり、様々なスケールでのモザイク状あるいはパッチ状の群集配置から構成されている。(露崎 2010)極相の有する3つの要因
極相種 (climax species)主に極相群集に出現する種のことを指す。極相に出現する種のことなので、極相群集をもとに作成される潜在自然植生の構成種が、おおむね極相種と一致する。ただし、多極相説で用いる極相の方が、単極相説よりも概念的に広いのにみられるように、極相種の範囲は厳密なものではないが、極相種には先駆種と比較して、おおむね以下のような特徴がある。1) 種子は、大きく、散布距離が短く、寿命が短い。2) 植物体は、成長速度が遅く、耐陰性が高く、寿命が長い。3) 繁殖は、開始時期が遅く、有性繁殖への資源投資が少ない。日本で極相種とみなされる高木種は、温暖帯では、シイ・タブ・イスノキなどが、温帯ではブナ・ミズナラ・サワグルミなどが、冷温帯ではオオシラビソ・トドマツなどがあげられる。これらの極相と認識できる生態系に特徴的にみられる動物種も、極相種と呼べる。(露崎 2010)極相群集 (climax community or vegetation) = 極相群落、極相植生極相に達したとみなされる群集を指す言葉である。ただし、極相概念が、気候的極相説、土壌的極相説などの説により定義が異なり、いずれの群集を極相群集と呼ぶかは立場により多少異なる。いずれにしても、極相群集は、群集の高い安定性と永続性をもとに認識できるとする点が、多くの極相概念間で一致している。ただし、安定性(stability)・永続性(persistence)の定義にも様々なものがある。一方、このことは、長期間維持されている群集である極相は、寿命や撹乱による様々な規模での部分的な群集構造の変化とその後の再生過程を経て、動的に維持されていることを意味する。A.S. Watt (1947)が提案した循環遷移(cyclic succession)は、群集が、さまざまな撹乱に応じた異なる発達段階の群集がモザイク状に入り組んだ形で成立する、という遷移観で、極相にもこれがあてはまるといえる。日本における代表的な極相群集は、森林となり、極相群集が、森林であることを強調するときに、特に極相林と呼ぶこともある。(露崎 2010)極相林 (climax forest)極相群集の一形態で、特に、森林であることを強調する場合に極相林と呼ぶ。森林は、通常、降水量が樹木成長に適量以上である地域に成立するため、日本のように降水量が豊富な地域では、極相は森林となるのが普通で、おおむね極相群集と極相林が同意となる。ただし、気候的極相説と土壌的極相説では、極相概念が異なるため、極相林が何を指すかについて、見解が異なるが、現在では、土壌的極相に相当する群集を含めて極相林ということが多い。日本では、南から北への緯度傾度に沿って、①暖温帯域の、タブ・カシ類・シイ類などの常緑広葉樹林または照葉樹林、②温帯域の、ブナ林などの落葉広葉樹林、③冷温帯域の針広混交林とエゾマツ、トドマツに代表される針葉樹林が、潜在的には極相林と呼べる群集となる。しかし、林相的には、これらの森林に区分できる群集でも、日本では、人為の影響を強く受けているところも多く、極相林と呼べる群集やそれらが存在する地域は少ない。(露崎 2010)植生が極相林の性格を持つ(推定) →
桜島 ≈ 1000年(田川 1964) 気候(的)極相 (climatic climax) (= 単極相, =mono-climax)F.E. Clementsが提唱した、遷移初期にはどのような群集であろうとも、最終的に1つの気候帯の中では唯一の極相に到達するという、気候的極相説にもとづく極相を指す。すなわち、極相は気候のみにより決められるとした極相観である。極相以外の植物群集は、すべて極相に向かい推移しているか、あるいは何らかの要因により推移が停滞した状態(亜極相など)にあると考える。極相は、優占種群が存在し、それらの種の安定性と永続性により認識される。気候的極相は、地球規模では、おおむね気候帯をもとにした群集区分であるバイオームに相当する。しかし、この極相概念は、実証性に乏しく、その後提案された多極相説、極相パターン説などの方がより遷移現象を説明しやすいことから、あまり用いられることはない概念となった。なお、現在の日本の多くの群集は、何らかの形で撹乱を受けた二次的な群集であり、気候的極相に相当する群集は、ほとんど存在しない(→ 極相林)。(露崎 2010)土壌的極相 (edaphic climax)多極相説(poly-climax theory)にもとづく極相概念では、同一の気候帯に属する地域でも、気候以外の要因、すなわち、土壌や微気象の違いにより、異なる極相群集が発達すると考える。それらの極相の中でも土壌要因により、群集構造を規定されている極相を指す。例えば、気候的極相説からはブナ林が気候的極相と考えられる日本の温帯域においても、乾燥土壌ではアカマツ林が、過湿土壌ではサワグルミ林などが発達し長い時間定着することがある。気候的極相では、これらの森林は亜極相として認識されるが、多極相的立場では、これらは土壌的に安定した極相群集とみなす。ほかにも、過湿土壌状態や低温のために、高木が侵入できない状態が長期間継続している湿原や高山草原などは土壌的極相とみなせる。また、人為により長期間維持されているススキ草地などの群集も土壌的極相とみなせる。また、多極相説からは、土壌的極相のほかに、地形極相、生物極相など、さまざまな極相を想定することができる。(露崎 2010) |
環境傾度分析 (environmental gradient analysis)(=環境勾配分析、傾度分析, = Gradient analysis) 群集が標高差や土壌の乾湿という様々な環境傾度に沿った配置様式を表現する分析方法全般を指す(s.l.)。環境傾度分析は、直接法と間接法の2つに大別される。直接環境傾度分析は、得られた環境データを分析に組み込み、環境と調査された調査区内の種組成との対応関係を調べる。一方、間接環境傾度分析は、調査された調査区や群集を、環境要因なしに種と調査区の配列を決め、ついで配列に関係する環境を抽出する。間接法には、主成分分析、(傾向化除去)対応分析などがある。狭義では、R.H. Whittaker (1967)が考案した、環境データと群集データの対応付け方法を指し、これは直接環境傾度分析に属する。正準対応分析(canonical correspondence analysis)は、環境要因を説明変数とした重回帰分析と対応分析を組み合わせた序列化手法なため、直接環境傾度分析に分類されることが多い。野外においては、必ずしも群集配置をうまく説明できる環境要因を抽出できるとは限らないので、直接・間接環傾度度分析は、適宜使い分けるべきである。(露崎 2010)極相パターン説 (climax pattern theory)環境傾度に沿い群集は連続して配置されているという植生連続説(vegetation continuum concept)に基づく、極相のとらえ方で、極相は、様々な環境傾度に沿って空間的に連続した各種の個体群の重なりあいのパターンとして認識できるとする説を指す。すなわち、時間的・空間的にもっとも成熟した段階に出現する種の組み合わせにより極相は形成される。Whittakerは、群集間には明瞭な境界が存在せず、エコトーンが幅広く存在する証拠を示すことで、この結論に到達した。この説によれば、極相的な群集は以下のように整理される。1) 群集の生産力と構造、個体群組成が安定した状態にある。2) ただし、群集は、環境傾度に対応した個体群の組み合わせのバランスから形成されているものなので、必ずしも群集や極相の存在を認識する必要はない。極相パターン説を証明するために、Whittakerらは環境傾度分析を導入し、環境と植物群集の対応関係を示す解析手法が大きく発展した。(露崎 2010)遷移相極相 (seral climax)変則遷移 deflected succession: 遷移系列の一部が欠落した遷移遷移途中で何らかの環境抵抗 → 途中で進行が止まった一連のclimax状態 これらは安定群集stable communityか? = climax → Clementsのmonoclimax theoryに対する反論 Ex. 山の斜面: 方角により温度等に差がありmesic stableになれずpreclimaxを形成。また、paleoclimatic (or postclimax) conditionを反映した植生relictとなっていることもある → 様々なclimaxが存在し、決してclimateがclimaxを決定するのではない 退行遷移 retrogression or retrogressive succession: 裸地化(遷移初期相)に向かう遷移 Ex. 過放牧地亜極相 (subclimax) (≈ 準安定相、妨害極相)気候的極相概念中で使用され、極相となりうる手前の状態で安定し、長時間にわたり遷移が停滞しているようにみえる遷移系列上の群集を指す(☛ 遷移)。極相に近づくにつれ、一般に遷移速度は遅くなるため、亜極相群集は長時間存在することとなり、準安定相とも呼ばれる。しかし、亜極相と極相の識別は概念的なものであって、実際に亜極相・極相を区分することは困難なことも多く、区分自体の意味も不明なことが多く、現在の生態学では、気候的極相概念にもとづく議論でもない限り用いない。極相に至らずに停滞する要因としては、噴火、山火事、人為などの撹乱があげられる。人為撹乱としては、ボタ山(ズリ山)や埋立地などが長期間経過し、草本期が持続すると、亜極相的とみなせることがある。これらの撹乱を除去すると遷移は進行するため、その意味で妨害極相disclimaxと呼ぶこともある。なお、前極相は、撹乱によらない要因により遷移が気候的極相の前の段階に留まることを言う。(露崎 2010)
準極相 proclimax: 亜極相は途中相を含む(批判) → 途中相除く亜極相 前極相 (preclimax) (=前安定相)気候的極相の考え方にもとづく概念で、気候極相となるよりも前の段階で安定している群集を指す。例えば、微気象などの差異で、本来、極相は森林となるべき地域で発達している低木林などが前極相に相当する。気候が同一とみなされる地域では、より乾燥した状態で成立する群集が、より湿性な群集に対して、前極相と呼ばれることが多い。しかし、この用語は、気候的極相説においてのみ使用する概念であるため、現在では、ほとんど使われない。(露崎 2010)後極相 (postclimax) (=後安定相)気候的極相説の立場で用いられる用語で、気候的極相よりも発達あるいは進展した状態で成立している群集をいう。たとえば、本来の気候的極相は草本群集であるにもかかわらず、谷地形である等の理由で土壌水分が増し低木や高木が侵入し、草本群集よりも発達したようにみえる群集を形成しているような場合をさす。しかし、気候的極相説においてのみ使用される用語であり、現在は、ほとんど使われない。(露崎 2010)単極相説 monoclimax theory気候によってclimaxは決定される (Climax is determined by climate)。climaxへの到達には十分な時間が必要(To be climax, sufficient time is required) → Whittakerのbiome概念
極相(climax): final stage
↑ 気候 → 温度: 年平均気温 最暖(寒)月平均気温
積算温度(吉良)等
降水量(年降水量)
↓ 降水様式 ↓ 植生の種類
植生の種類 → 光 気温
O2 温度 → 光合成エネルギー 有機物
CO2 水 ↓
生物 → 土 → 無機物
(バクテリア・菌類・藻類)
多極相説 polyclimax theoryEx. エゾマツ・トドマツ = Climatic climax species
シラビソ.オオシラビソ.エゾマツ = 土壌発達 ↔ → 多極相説: 極相は1つに限定する必要ない 気候 + 土壌(土壌極相)(Ex. アルカリ土・塩性土・石灰土・蛇紋岩土) + 人為干渉(妨害極相) 偏向的極相 plagioclimax: 動物(人間)の作用等の原因が働いて極相へ向かうコースからずれたもの |
遷移モデル
|
定着促進効果と競争 facilitation vs competitionDef. 定着促進効果: ある生物種が他種の定着・生育・繁殖を助ける作用
撹乱地やストレス環境下でみられる ☛ 駒ヶ岳のミネヤナギ = 共栄植物・共生植物・共生植物 Ex. シソ(コンパニオン): 害虫忌避・生育促進 → ナス (京都伝統野菜畑) ストレス勾配仮説 stress gradient hypothesis一次遷移 ----- 二次遷移 ----- 極相
阻害____< < < < < < < < < < < < < < < < < |
タストロフ分岐点(catastrophic bifurcation)これを超えるとある安定点から別の安定点に急速で不連続な変化を起こす点
サハラ砂漠: 急激な砂漠化 |
履歴現象 (hysteresis)ある量Aの変化 → 他の量Bが変化Aの変化量は同じでも変化経路によりBの値が異なる現象 |
|
(植物)群集において、その構造と種構成が時間に応じ変化する一連の組合せ (「時系列」は統計学用語で既にあるので、そう訳せないことが悲しい) Ex. 森林火災、氷河後退、溶岩流出、スキー場造成 - それぞれの撹乱発生時期が明瞭なところでの時間系列に沿った群集の変化 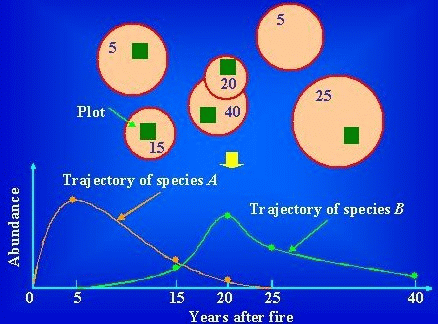 図. 時間系列アプローチによって遷移系列を得る仮想的な手順。これは、火災後の調査区の場合を示している。
図. 時間系列アプローチによって遷移系列を得る仮想的な手順。これは、火災後の調査区の場合を示している。[上図] 円は、火災を受けた範囲を示す。四角は、森林火災区域内に設定された調査区を示す。数字は、火災からの経過年数を示す。[下図] 種構成の連続変化。種Aは火災から5年後に優占し、ついで次第に優占度を減少させる。種Bは、火災から20年経過したときに最も優占する。仮に、時間の経過につれ種構成が変化するとすれば、種Aが、その地域の火災後の先駆種と特徴づけることができる。 |
Ex. 桜島溶岩 Ex. スキー場斜面 (Tsuyuzaki 2005) 時間系列の仮定に関する問題調査区間の撹乱の強度・規模・頻度は常に一定とは限らないが、そう仮定せざるをえない。傾向を見るのには良いかも知れないが、細部を知ることはできない |