(Upload on February 17 2026) [ 日本語 | English ]
Mount Usu / Sarobetsu post-mined peatland
From left: Crater basin in 1986 and 2006. Cottongrass / Daylily
HOME > Lecture catalog / Research summary > Glossary > Seed dispersal
[evolution, succession, seed dispersal ( animal ), seed trap ]
[seed germination, mast seeding (豊凶), seedbank (estimation methods, seed dormancy]
The passive or active movement, usually one way and on any time scale, of organisms from their point of origin to another location where they may subsequently produce offsprings (e.g., insect larvae, fungal spores, or plant seed by wind; insect adults by flight; animals by walking or swimming).
Primary vs secondary dispersal (一次・二次種子散布)Primary seed dispersal (一次種子分散)That means seeds dispersed by any dispersers, such as wind, animal, and gravity.Secondary seed dispersal (二次種子分散)The secondary dispersal occurs when seeds are moved elsewhre after the primary seed dispersal is completed.Evolution
|
Suceession (遷移)In the early stages of succession, e.g., soon after volcanic eruptions, species with long-distance seed dispersal, such as wind-dispersal, immigrate faster. |
|
Table. Seed dispersal types Dispersal vector (term): species
*: species producing fine, such as Drosera spp. seeds are considered to be wind dispersal. |
Classification based on the transporation meansectozoochre
epizoochores (exozoochores): on surface such as skin
endozoochores: through body inside Classification based on the vectors
mammaliochory: mammal
Higashi et al. 1989. Adaptive advantages of ant-dispersed seeds in the myrmecochorous plant Trillium tschonoskii (Liliaceae). Oikos 54: 389-394 |
Trillium tschonoskii Maxim. is a myrmecochorous perennial herb which grows in single-species stands in the cool temperate broad-leaved deciduous woodland of Hokkaido, northern Japan. Its many-seeded fruit initially falls close to the parent, over 50% within 20 cm. Ants, principally Myrmica ruginodis and Aphaenogaster japonica, transport seeds to their nests, a mean distance of 64 cm. The nests are overdispersed and short-lived, and are no richer in nitrogen or phosphorus than surrounding soils. There are proportionally more older (3-leaved) than younger (1-leaved) juvenile Trillium plants at distances > 60 cm from the nearest parent plant than at 0-30 or 30-60 cm. This indicates that the relatively short-distance dispersal of T. tschonoskii seeds by ants is sufficient to reduce seedling mortality, by reducing competition between seedlings; and that this is the primary advantage of myrmecochory in this species. This contrasts with previous studies, involving ant species with longer-lived nests, where the main advantage to the plant is nutrient-enhanced seedling microsites. saurochory: reptileichthyochory: fish 
Cache (隠匿散布)Seed cache is a kind of animal dispersals.Although animals represented by rodents and birds that cache acorns under the ground surface, they often abandon to eat. When the seeds are left there, they can germinate and this means seed dispersed by animals. |
Importance of uisng seed trapsTo estimate what species disperse how many seeds to where, we have to capture seeds in various locations. To capture seeds effectively, we have to develop more precise traps. There are two types of seed traps, dry and wet types. Every seed trap has advantages and disadvantages. Therefore, when we decide what types of seed traps we use, we must consider what is our major interest in the study. Seed traps are often used as litter traps.
Dry-typed seed trapForest (widely used)This is a popular tool for collecting seeds in less-windy ecosystems, such as well-delveoped forests. The target seeds were gravity-dispersal seeds, e.g., acorns, and this type works well. However, light seeds, such as wind-dispersed seeds, escape from the traps. To increase seed recovery, we have to invent more excellent traps.→ in a forest on Mt. Makiling, Los Banos, Philippines, on December 17 1997 (Taguiam & Tsuyuzaki 1998). 
Alaskan forestThe surface to capture seeds is made of artificial lawn. To capture Picea mariana seeds, this trap is designed. The four corners of tray are fixed with the ground surface by long nails.→ close-up of a trap. On May 10 2006 at Poker Flat after the 2004 wildfire (森林火災) (Tsuyuzaki et al. 2014). |
Wet-typed seed trapTo avoid seeds that escape from the inside of traps, sticky materials are paseted on the surface of traps. To increase seed recovery, we have to change seed traps frequently, in particular, when we set them up in dusty habitats.
 [1] seed traps set up in an abondoned peat-mined site on Sarotetsu wetland by A. Koyama. The traps were set up on the center, edge and flat of tusscoks. [2] the trap captured many seeds of Eriohoprum vaginatum. [1/2] on July 5 2007. (Koyama & Tsuyuzaki 2010)
[1] seed traps set up in an abondoned peat-mined site on Sarotetsu wetland by A. Koyama. The traps were set up on the center, edge and flat of tusscoks. [2] the trap captured many seeds of Eriohoprum vaginatum. [1/2] on July 5 2007. (Koyama & Tsuyuzaki 2010)
|
|
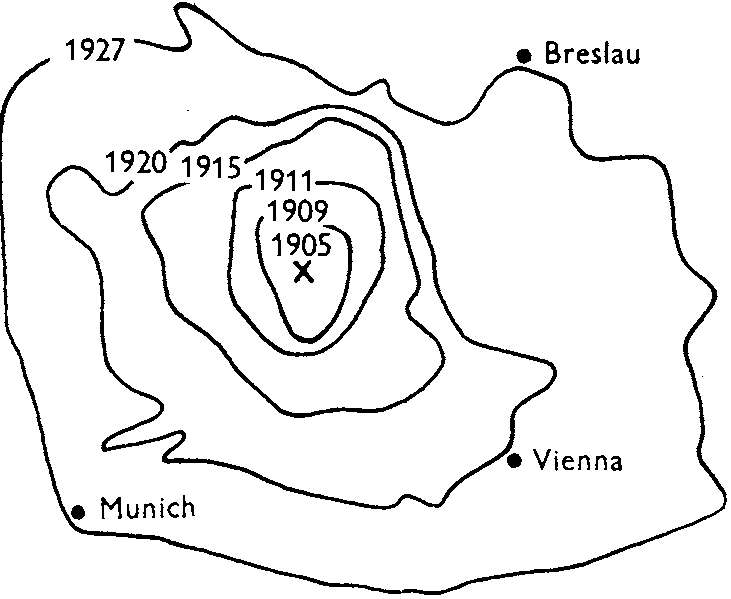
Def. Invasion (侵入): the process by which non-native species immigrate an ecosystem and establish the populations Def. Propagation (伝播): the process by which organisms reproduce and spread throughout their environment 1951 Skellam JG (1914-1979): invasion biology (reaction-diffusion model)
Ondatra zibethica L. (muskrat)
→ spread throughout Europe 50 years later
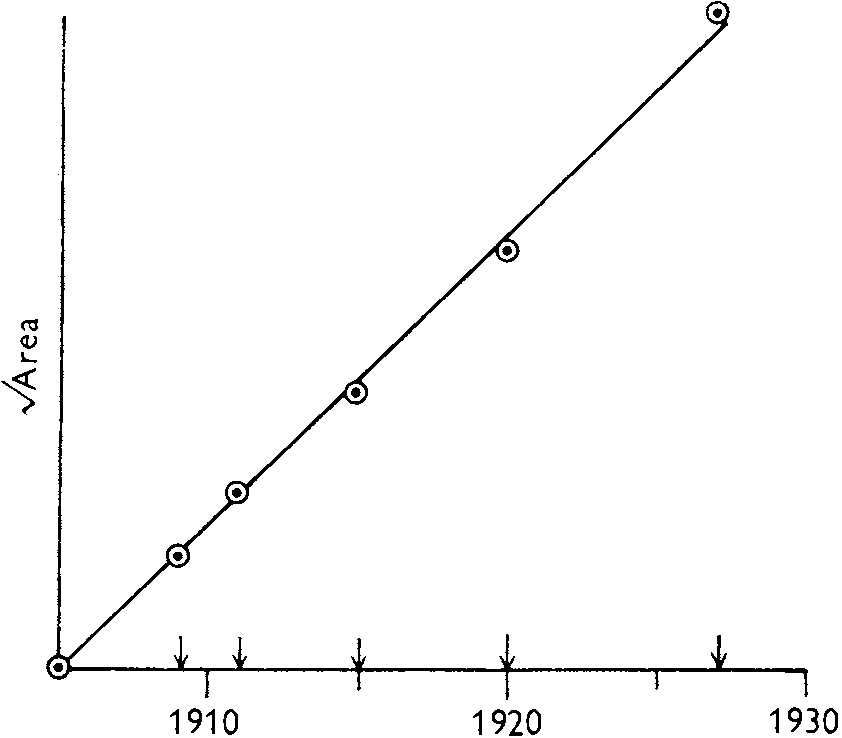
migrated using traffic networks area at initial stage: 145t2 → later: exp(0.35t) curve at initial stage = π(ct)2 → c = 0.13/yr
(a) λ0 = 0.13/yr. (b) λ1 = 2 × 10-3/km/yr. (c) λ2 = 2 × 10-5/km2/yr |
1988 Lubina & Levin: Enhydra lutris in California
1914: considered to be extinct - rediscovered on Sur Cape conservation - reproducing and spreading well Before 1972: 1.4 km/yr to north. 3.1 km/yr to south reached to Monterey Bay and Cayucos Cape - boundary After 1972: accelerated 1988 Okubo: Sturnus vulgaris in USA
first 8 years: stagnant spread → 15 yrs: slow spread → accelerated c = 11 km/yr, L = 300-400 km, λ1 ≈ 0.02/km/yr other birds, e.g., Passer domesticus and Carpodacus mexicanus, showing similar spread patterns 1989 Hengeveld: Cervus elaphus invaded to New ZealandThe spread pattern is not linear with time, unlike Fig. 2, probably because the area is limited on islands
1851 firstly introduced C. elaphus in the South Island
firstly discovered from Japan in 1976 at Nagoya Invasion of competitive speciesEx. invasive Sciurus carolinensis vs native S. vulgaris (Okubo et al. 1989)ε1 = 0.61/yr. ε2 = 0.82/yr ε1/μ11 = 0.75/ha. ε2/μ22 = 10/ha empirically μ12 > μ22 μ11 > μ12 S. carolinensis expands with the exclusion of S. vulgaris ε1/ε2 = 0.74 and μ21/μ11 < 1 ∴ c~2 ≈ 2√(ε2D2) = 2√(0.82D2) empirically c~2 = 7.7 km/yr ∴ D2 = 18 km2/yr |