(2016年12月10日更新) [ 日本語 | English ]
HOME > 講義・実習・演習一覧 / 研究概要 > 小辞典 > 生態系
[ 群集 | 環境 | 生態系 | 保全 | 生態系サービス ]
|
= 共同体 (≈ 群落) → 異種個体群集団
Ex. プランクトン群集 (注: 群集単位 = 群集 association = 植物社会学用語) Ex. 森林や草原 = 植物群集 + 森林中の動物 + 土壌中小動物や微生物 植物以外も含め生物群集を扱うのが正しい ある土地に生育する生物の集団を全体的に漠然と指す (s.l.)ある環境内で一定のまとまり保つ生物個体群の集まり (s.s., Humboldt 1805) 各種個体群は有機的相互関係を有する 優占種を基に2群集認識可 ↔ 連続変化とみる(Whittaker説)
群集間相互作用 = 収聯 (interactions between communities will be converge) ニッチェ(生態的地位) (ecological) niche生物群集を構成している個体群には、食物連鎖における位置と群集内部で占める空間的位置がある。この2つの位置のこと。各々の個体群にとって生活のエネルギーと住む場所の獲得を意味している生物類縁関係 = classification, ecological niche。全く異なる生物群集に類似点が存在 – 系統進化 phylogenic evolution: taxonomyの上全く異なる種でも同じnicheを占めることはある |
1927 Elton: 群集内におけるある生物の地位 status 1954 Clarke 機能的ニッチ functional niche + 場所的ニッチ place niche 1957 Hutchinson:
群集側から見ることを一先ず捨て、あらゆる環境条件に対する個体群の反応をn次元に展開した多次元地位概念提唱。ニッチは物理環境だけでは決定されない – 競争 ニッチ幅 niche breadth or niche width (van Valen & Grant 1970) ニッチ空間中の有る特別な線に沿った距離をもととしたニッチの幅 ニッチ重複 niche overlapEx. オーストラリアにおけるトカゲが捕食するシロアリ
トカゲ(体重) 捕食餌(シロアリ)サイズ |
|
植物群落(群集) plant community: 食物連鎖の出発点 = 生物群群集の性格を表わす面強い ⇔ 動物群集 植物群集とは非常に幅広い言葉 → バイオーム (biome), 生物圏 (biosphere) 植生 plant community (vegetation)ある土地に生育している植物の集団を全体的に漠然と指す (s.l.)
講義では、1の意味でのみ使用している。 植生構造評価基準 (criteria of vegetation structure) |
相観·構造·生活形·優占種·種組成·機能·遷移過程·生育地等の特徴によって色々な把握の仕方ができる(それでいいのか)
相観 physiognomy優占種生活形: 生物群集の特徴を視覚的にとらえたもの = 「見た目」最上層構成植物形を用いること多 (Ex. 落葉常緑樹林)。個体を指すこともある 代表的な基準
|
植物群集とは現実かまやかしか → 植物群落認識論
| 提唱者 | Clements | Whittaker |
| 説 | 単一極相説 climatic climax hypothesis | 植生連続体説 individualistic hypothesis |
| 種間相互作用 species interactions | 強い strong | 弱い weak |
| 種パターン species patterns | 具体的に変化 change in concrete | 徐々に変化 patterns change gradually |
| エコトーン ecotone | ない | 一般に存在 |
| アソシエーション association | 具体的 | 抽象的(概念的) |
| 歴史的背景 (植生概念相違) | ↓ | ↓ |
| アプローチ approach | 分類法(植生単位観) ZM - 植物 ⇔ 場所分類法 = ヒエラルキー系 英豪 - 植物 → 場所分類法= 網目系 |
序列法(植生連続体観) 合州国 植物 → 場所序列法 (連続体分析·比較序列法) 立地 → 場所序列法(環境傾度分析) |
[植生の垂直分布]
環境の構造帯状の概念zonal concept-空間的概念(秩序)温度・湿潤度 Ex. Köppenの気候区分, 垂直分布 → 配列 zone 帯状zonal: 成帯性土壌 zonal soil、気候的極相等
非帯状 azonal: zonalでない部分が比較的認められるもの |
= 風衝・土壌・水分等で極地的に形成されたzone 超帯状性 suprazonal: 人工的なものに多い極めて明瞭な帯状 エコトーン ecotone2つの植生の間(境界)に形成された2植生とは異なる植生
草原 - 低木林 - 森林 移行帯または推移帯(訳がよくない)とも呼ばれ、陸域と水域の境界になる水際のことをいう。 - エコトーンと推移帯は違う |
|
= Umgebung, Umvelt 環境 environment: Σf: abstract or generalized concept - 抽象的 (f: factor) 環境要因 = 無機的環境要因 + 生物的環境要因: 種々環境要因は統合され、一体となり生物に働く 生息地(生育地、産地、立地, habitat): concrete or specific environment - 現実的・具体的 クロード・ベルナール(生理学創始者)が内部環境と外部環境を分ける内部環境: 細胞の間を満たす体液の状態 維持 - ホメオスタシス 外部環境 限定要因 limiting factor 要因群中、生物が必要なもので最も不足(過剰)したで生命活動が著しい影響を受ける リービッヒの最少律(最少量): 成長速度 ブラックマンの限定要因説: 光合成速度 緑色植物の光合成 → 環境要因 = 光・温度条件
環境要因中、1つでも不足したり、逆に過剰になると、生物の営みが抑制されることがある 作用・反作用・相互作用作用 action: 環境 → 生物反作用 reaction: 生物 → 環境 相互作用 coaction: 生物 ↔ 生物 時間的・空間的にからまり複雑化 → 遷移 succession はこれらの産物 |
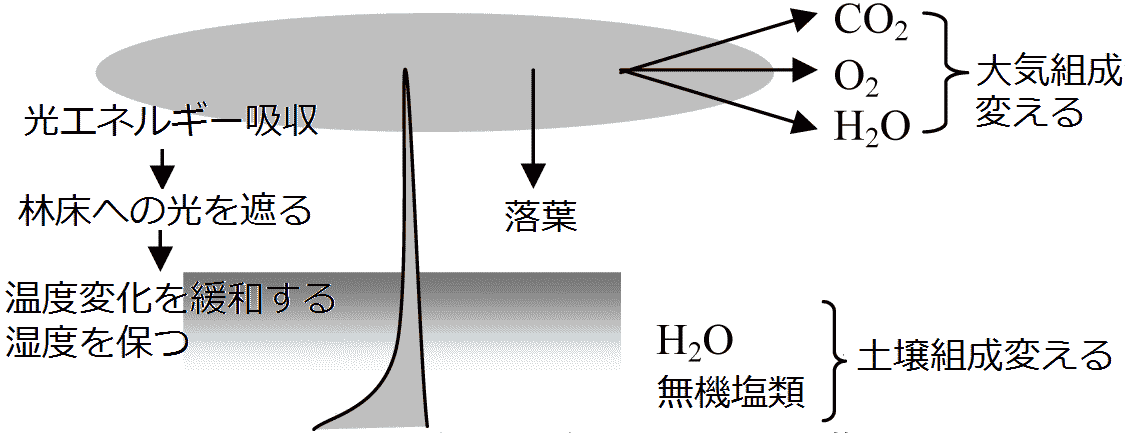 図. 樹木の環境形成作用(反作用) Ex. ダーウィン: ミミズの土壌に対する反作用研究 → ミミズは落葉の混じった土を食べ1日排糞量は自分の体重とほぼ等しい。イギリスのある牧草地では10年間にミミズが積み上げる土壌の厚さは2.5-3.8 cmに達する 作用-反作用系 action-reaction system (穂積 1973): 生物 → 環境変化
環境脆弱性指数 (environmental vulnerability index, EVI)2004年開発終了脆弱性: あるシステムの特性が極限的事象のインパクトを最小化あるいは吸収する潜在性 50の指標の組み合わせ
危険性 32 ⇔ 経済面・社会面の脆弱性指数とは性質が異なる |
|
Tansely (1935)提唱: ある地域に住む全ての生物と非生物的環境を一纏めにし、主に物質・エネルギー循環に注目し機能系として捉えたもの(Odum 1971, 1989)。情報を第3のフローとして加えることがある 生態系 = 植物群集 + 動物群集 + 環境生態系概念は広く、また、そうあるべき(broad concept) (Odum 1971)
必然的関係 → 機能的統一体 functionally integrated system 生態系機能 (ecosystem function)生態系内相互作用による物質の生産、分解、循環に代表されるプロセス
Ex. 光合成 (photosynthesis) ヒトのエネルギー代謝量(環境への反作用)生物エネルギー消費: 他生物利用できないエネルギー(電気、熱、化学等)を社会的生活エネルギー源として利用→ ヒトは地球のエネルギー循環に強力な影響を与え、時にはエネルギーや物質循環コースを変更
表. 人間1人1日当たり物質収支 (Talbot 1965) (数字は変わってるはず)
vs 生物としてのヒト(食料) = 15000 cal/g/yr → 13:1 |
生態系の寿命, T開放系(資源有限) Ex. ルーメン lumenT = M/(B·m)
M: 資源貯蔵量 寿命延ばす → M↑, B↓, m↓ 開放系(再生循環系あり)T = (M + Mo)/(B·m - Br·mr)
Mo: 再生循環する物質量
i) Mo = 0, Br·mr = 0 ⇒ 開放系 メタ群集 (metacommunity) |
静的モデル → (タイムラグ考慮) → 動的モデル
不可変モデル unchangeable model= 静的モデル static system model時間軸を考慮しない場合このモデルはある程度適用可能 Ex. 人口問題: population unit: age structure - その時点での構造を調べる = age structureの不均衡 1) 繁殖を推進するようなエネルギー物質の供給 2) 死亡率を最少化するような好条件の成立 可変モデル changeable model= 動的モデル dynamical system model変化の受容体receptor又は供給体は複雑化する 1) 下位クラスは上位クラスに包括される 2) それぞれのクラスは全て相互作用で結ばれている |
定常開放系 (steady-state open system)= 散逸構造 (dissipative structure)熱力学的に平衡でない状態にある開放系 エントロピー増加分を外部に捨てることのできるシステム
地球: 大気圏内の水循環と対流を通じて熱エントロピーを圏外に捨てている 生物地球化学モデル物理的環境因子(気温・降水量等)を用い、植生構造と機能の変化を予測するシミュレーションモデル |