(2023年9月21日更新) [ 日本語 | English ]
HOME > 講義・実習・演習一覧 / 研究概要 > 小辞典 > 光合成
|
[lE ≡ 光エネルギー] 生命は光によって空気から織り出される緑色植物(s.s.)と光合成細菌(s.l.)がlEを利用しCO2と水から有機物を合成する過程= 植物によりlEが生物利用可能な自由エネルギーになる変換過程 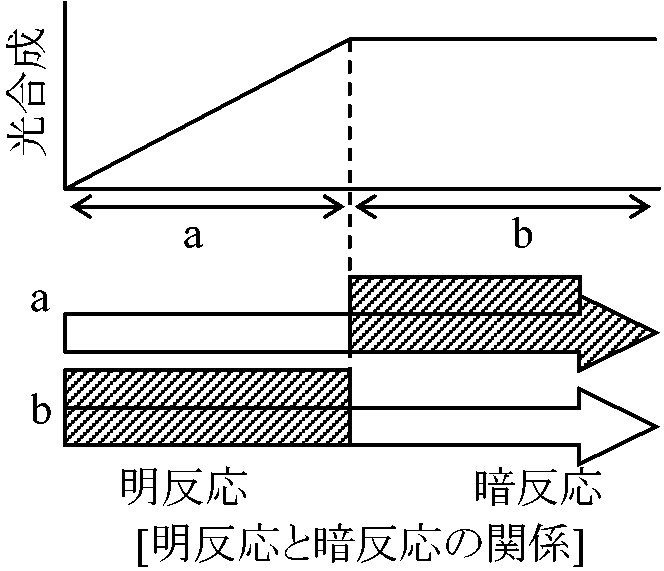 [2反応] 光化学反応(明反応) → 化学反応(暗反応)
[2反応] 光化学反応(明反応) → 化学反応(暗反応)明反応 light reaction: lE吸収し(= 光必要)、そのエネルギーを電子伝達と光リン酸化エネルギーに変換する反応 暗反応 dark reaction: 明反応後に起こる無光状態でも起こる反応。NADPHの還元力とATPエネルギーは、CO2固定回路であるカルビン回路によりCO2還元に使われる 緑色植物(C3)の光合成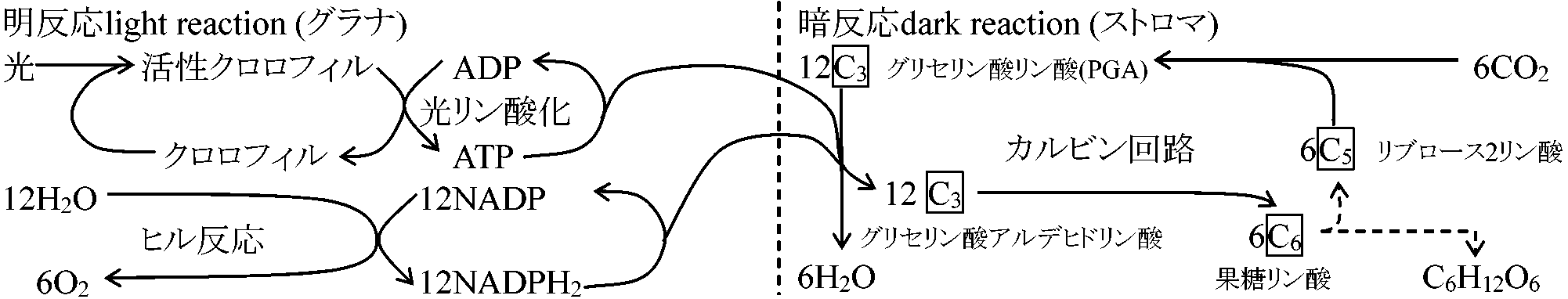
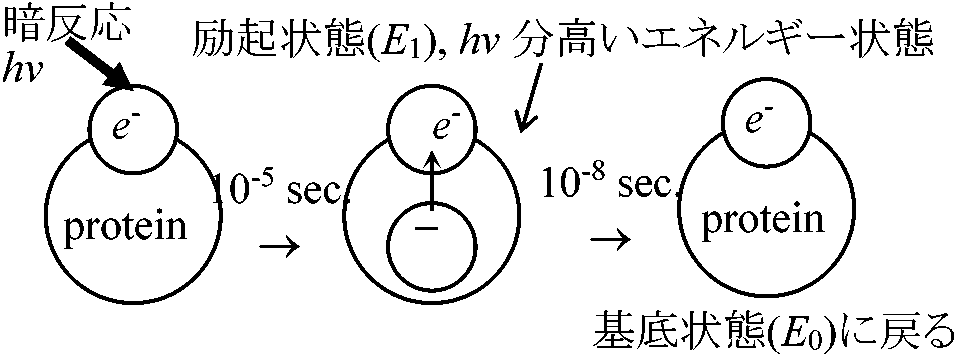
明反応部クロロフィル(Chl)吸収lEによりH2O分解されH(NADPと結合し暗反応へ)とO(気孔から体外放出)になり、同じくlEによりATP(暗反応に使われる)が合成される。光強度と波長が関係し温度にほとんど無関係1) ChlがlEを吸収し活性化され高エネルギーの電子を含む活性Chlになる 2) 活性Chl中のエネルギーによりH2O分解されO2放出 3) ヒル反応で発生したHは葉緑体中の有機化合物と次々に結合分離しつつ受け渡され、最後にNADPに移され、この物質をNADPH2に変える 4) HがNADPH2に渡される反応の一方で活性Chl中のエネルギーの一部を使いATPが形成される(光リン酸化反応)。生じたATPは暗反応に必要なエネルギーを供給するのに使われる 暗反応部暗反応で生じたH2とATPの働きでCO2が還元されブドウ糖に同化される。[カルビン回路を含む熱化学反応(ブラックマン発見) = ブラックマン反応ともいう] CO2濃度と温度が関係し光は不用1) CO2がリブロース2リン酸(C3)に取りこまれて2分子のグリセリン酸リン酸(C3)になる 2) グリセリン酸リン酸は明反応で生じたNADPH2とATPの働きにより還元されてグリセルアルデヒドリン酸(C3)になる 3) この3炭糖リン酸は2分子づつ結合して果糖リン酸になり、その一部がリン酸を離してブドウ糖になる 4) 一端CO2と結合して消費されたリブロース2リン酸は複雑な中間生成物を経て再合成される 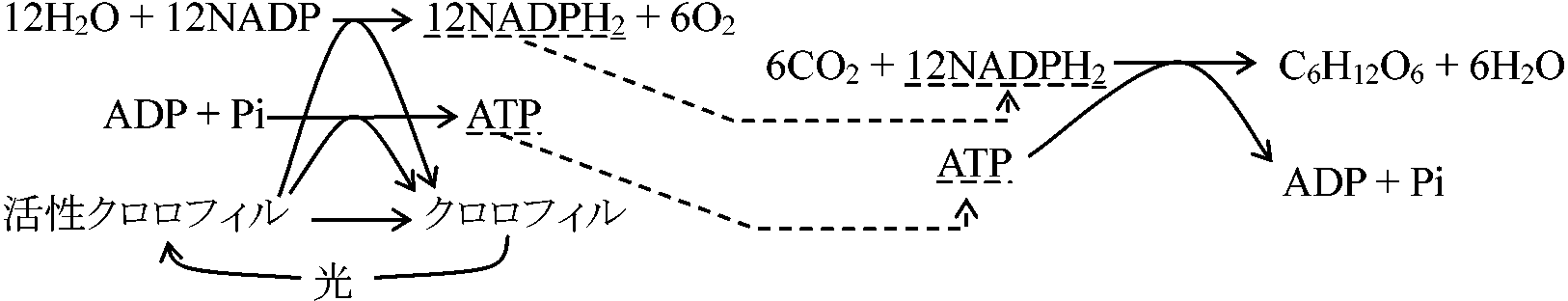 6H2O + 12H2O + 688 kcal → C6H12O2 + 6H2O + 6O2 6CO2 → [18ATP + 12NADPH2] → C6H12O6 [1 mol CO2の還元に3 mol ATPと2 mol NADPH2が必要] 高等植物における光合成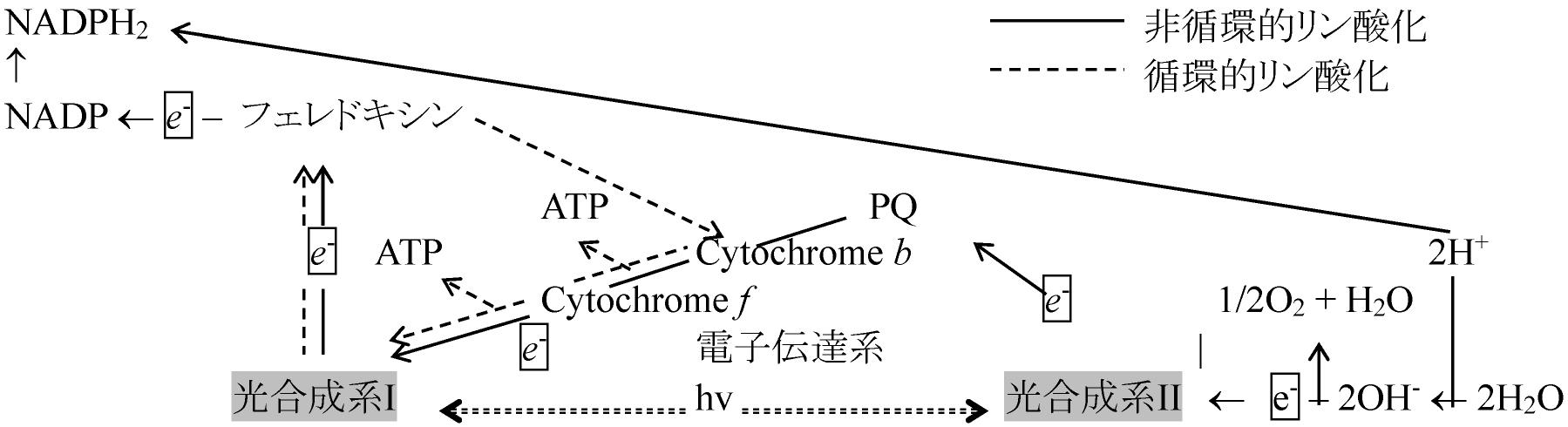 本来エネルギーが高順位になると状態量的に低順位に戻るが、光合成ではエネルギー(e-, 電子)がプラストキノン(PQ)に渡り、さらに種々チトクロームに渡り最終的にプラストシアニンに渡り、クロロフィルaにエネルギーが渡される。その途中過程で、徐々にエネルギーは外へ出てそのエネルギーによりATP生成される。光合成系は、発酵系を基に高次な光合成系へと進化してたものと考えられる NADH → FADH → CoQ(呼吸) → Cyt b → Cyt f … → Cyt a → … ATP _______________PQ (光合成) 光エネルギー solar energy表. 波長におけるエネルギー量: (赤色 = 低エネルギー) ↔ (青 = 高)
色________赤_________________________________青 |
光合成と関係する光 葉に当たった光
葉に当たった光
反射光 1883 Engelmann
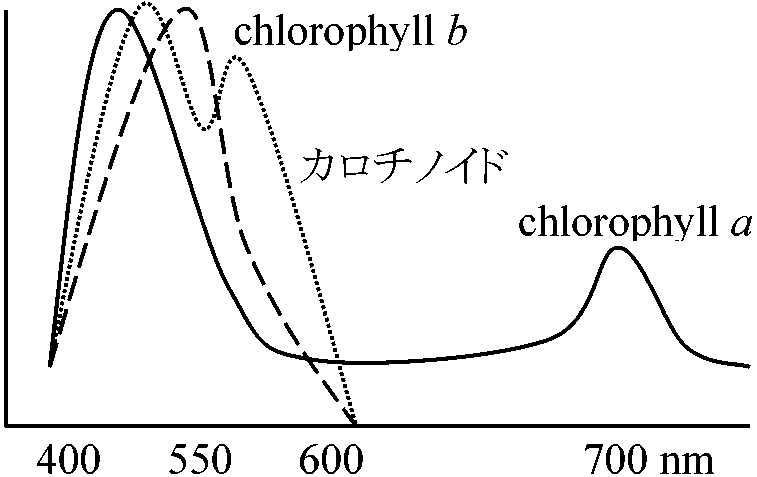
_____↙_____↙ 吸収スペクトル(利用光) P700 ≡ 700 nmに最大の吸収を持つ色素 P680 (P690, P700): 細胞に光を与えると色素色消失 → chl. a + proteins複合体complex 光量子としてのenergyを捕まえるのはchlorophyll aの一部分の分子だけである(0.1-0.003%) Chl. a → 葉緑体中と葉緑体内では吸収スペクトル異なる → 同じ色素でも存在様式が異なる ⇒ 微分スペクトル法開発 反応中心クロロフィルreaction central chlorophyllと集光クロロフィルlight harvesting chlorophyllが存在 反応中心クロロフィルは酸化還元を行う → チトクロームfと関連した反応中心
P700 or 680 → PS I (Photosynthesis I)の反応中心クロロフィル 光化学系 I (photosystem I, PSI)1939 Emerson: 光合成系I, PSI: 710 (700, 680) nm - 光合成起る活性クロロフィル(Chl) a(励起Chl)分子の電子を使いNADPH2にする。Chl aの電子はNADPに入り、その代わりにPS II(Chl b中心)から電子伝達系を経て電子供給されChl aは元の状態に戻る。活性Chl電子がNADP還元に使われないと電子伝達系に戻り電子伝達系を渡る時にATPが生成される 光化学系 II (photosystem II, PS II)670 (690, 640) nm - 光合成促進活性Chl aの電子放出で生じた酸化力により水を加水分解し酸素を発生する系。H2Oは2H+と2OH-に分解され2H+は最終的にPS Iの電子e-と結合しNADPH2作る。この際、電子伝達系でATP生成される。他方2OH-は電子を奪われ1/2O2とH2Oになり、その際の放出電子がPS IのChl aに渡され電子補充に使われる
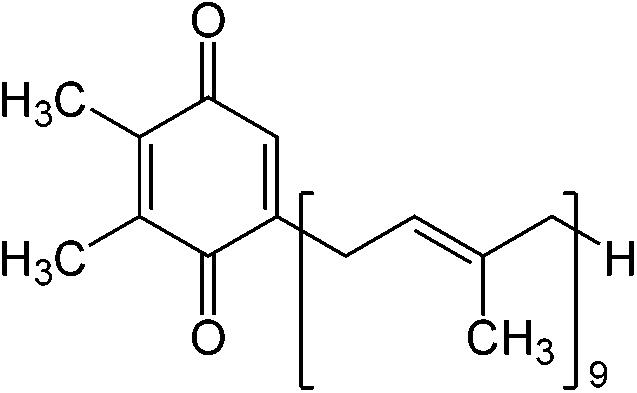
[chl. a: 200個/分子 + chl. b: 50個 + carothinoids: 50-200個] 還元力とATP生成集光クロロフィル → chlorophyll a, b, c, d, (e), bacterochlorophyllユビキノン ubiquinone cyclic electron transfer system: e- → Fd, NADH H+ → proton motive force → ATP生成 |
|
(Calvin & Benson 1957) カルビン(=ベンソン)回路 Calvin(-Benson) cycle= 還元的ペントースリン酸回路、光合成的炭素還元回路、C3回路14Cを用い炭素固定経路での産物の流れを明らかにする = Clorella, Scenedesumus (材料) グリセリン酸リン酸、リンゴ酸、コハク酸、アスパラギン酸、アラニン等を14Cラベル 葉緑体ストロマstroma部分にて行われる 光リン酸化 photophosphorylationクロロプラスト chloroplast____Step 1______↓ chlorophyll-protein = 明反応 ___________H2O → 2H+ + 2e- + 1/2O2 _______________↑ 光分解: light energy (to pigment chlorophyll) ____Step 2__NADP + 2H+ + 2e- → NADPH2 ____Step 3__________↓ 光リン酸化反応 ___________ADP + Pi → ATP クアンタゾームquantasome明反応の最少単位= 明反応可能な全要素が含まれる最小値 = グラナラメラ膜を切り明反応停止直前の大きさのグラナラメラ 高等植物においてchl. a:b = 70:30 or 75:25
2nH2O* + nCO2 → (CH2O)n + nH2O + nO*2↑ → 酸素は水起源 |
ADP + Pi ← 光
P680 (P680 + e-, e-の受取先はH2O) (Hill 1947) ヒル反応 (Hill reaction)1937 Hill R: spinach → isolated chloroplast: ferrie oralate(シュウ酸第2鉄)
CO2与えないが酸化される: 酸化 = (電子消失/水素消失/O獲得)
H218O → 18O2 |
エマーソン効果: 光合成に重要な青色(400-500 nm)・赤色(600-700 nm)に、近赤外線(700nm)以上の波長を加えると相乗効果で光合成が促進される効果 → 赤外線のみでは温かいだけで、光合成に効果はない
型 緑色植物型 細菌型
O2発生 伴う 伴わない
主要光合成色素 Chlorophyll a Bacterio-chlorophyll
利用光 赤色光 近赤外光
H-供与体 H2O H2O以外にHe, HS等
エマーソン効果 認められる 認められない
藍藻 (原核生物, prokaryote)光合成特別器官持たず、体全体にチラコイドを持ちそこで光合成を行う
CO2固定 → O2放出
hv____________ hv
Anabaena Anacystis Porphyridium
PS I 9 16 5
PS II 91 84 95
PS I 71
PS II -
細菌型光合成細菌は大部分が従属栄養生物だが、一部特殊な細菌が独立栄養生物1. 光合成細菌 2. 化学合成細菌 (別掲) 光合成細菌嫌気的に炭酸固定・O2発生なし[反応一般式] CO2 + 2H2A → [CH2O] + H2O + 2A Ex. A = H: [CO2 + 2H2O → CH2O + H2O + O2] 紅色イオウ細菌 chromatiaceaeA = × CO2 + 2H2S + 4H2O → [CH2O] + 1/2·H2SO4 + H2O 緑色イオウ細菌 chlorobiaceaeCO2 + 2H2S → [CH2O] + H2O + 2S 非イオウ紅色細菌 rhodospirillaceae
2CO2 + Na2S2O3 + 5H2O → 2[CH2O] + 2H2O + 2NaHSO4
Case. 非イオウ紅色細菌: |
還元的カルボン酸回路(還元的TCA回路): 一部の独立栄養細菌 1966 Evans, Buchanan, Aron: TCA回路の逆行 Ex. Chlorobium, Thermoproteus
還元型フェレドキシン(Fdred) を使いAcetyl-CoAとCO2からケト酸(α-KG)ができる2つの反応によりCO2を固定 → 2オキソグルタール酸 + Co-A-SH + Fdox CO2 1分子固定: (ATP 3 mol = 36 Kcal) + (NADPH2 2 mol = 104 Kcal) = Total 140 Kcal = 光エネルギー Chlorobium thisosulfatophilum: 高濃度CO2 - 還元的カルビン回路働く 化学合成細菌の炭酸固定無機物または有機物の酸化エネルギー(化学エネルギー)使い炭水化物作るカルビン回路で固定。PEP carboxylase関与不明。cytochromes関与は証明された 一般式: CO2 + 4[H] + Energy → [CH2O] + H2O
NAD(P)のE0' = -0.32V カルビン回路 → 主経路 / 赤色カルボン回路 → 少ない イオウ細菌 Thiobacillus
H2S + 1/2O2 → S + H2O_____________________50 Kcal
Nitrosomonas 亜硝酸菌: NH4+ + 1/2O2 → NH2OH + H+___3.9
温泉・土壌 Ex. Pseudomonas fracilis, Alcaligones eutropus, Paracoccus denitrificans CO2固定経路RuBisCo (Ru-1,5-P2 carboxylase): カルビン回路の酵素14CO2 → Products (カルビン回路で固定 - いずれもこの経路だけではない) Ex. PGAの代わりにglucose, malateが生産物として見つかる事例 Clostridium, Desulfovibrio: heterotrophic = 有機物, anaerobic = H2 |
|
同化箱 gas exchange system or photosynthesis measurement chamber
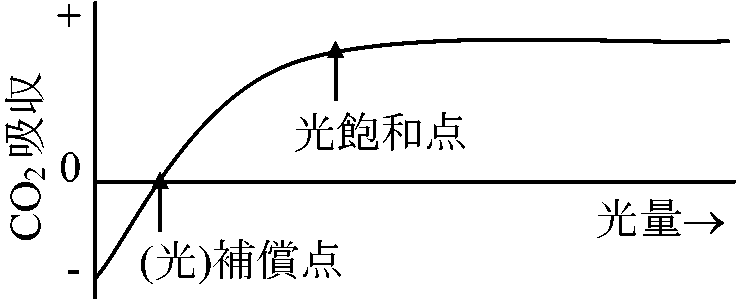
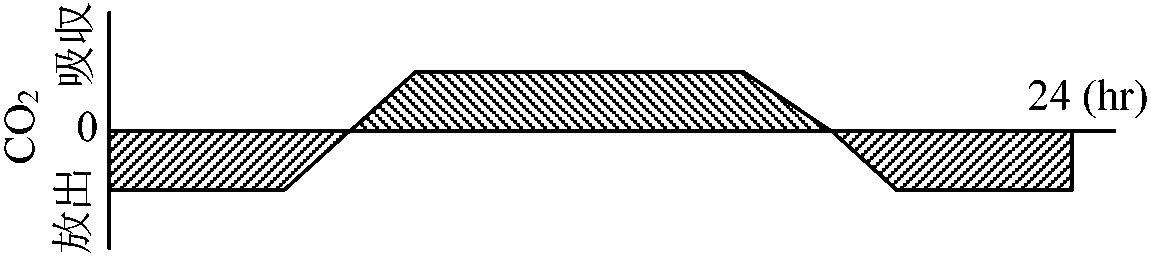
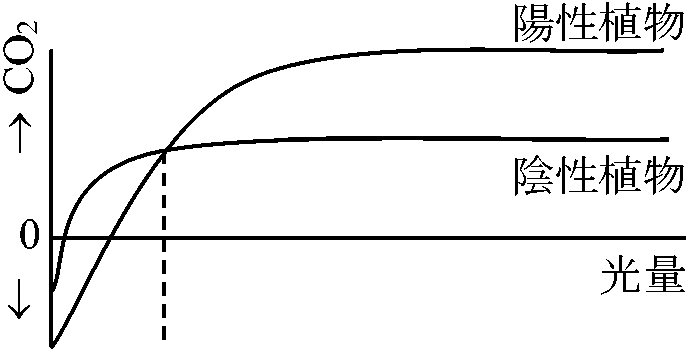
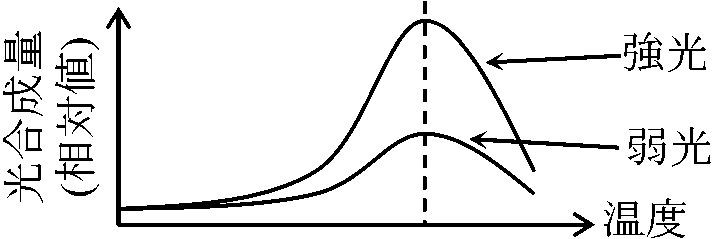
植物体の一部または全部を封じ込め同化(光合成)を測定する容器 光-光合成曲線(温度一定)高等植物における光合成組織・器官: 葉 = 葉緑体
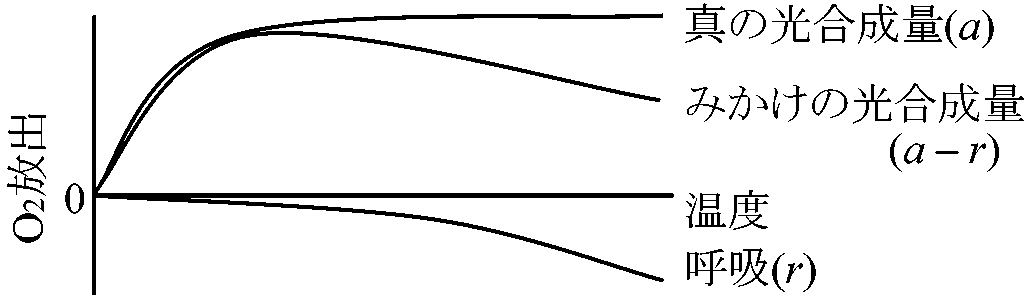
*: 水草実験ではCO2は水に溶けるためO2放出量を測る (光)補償点: みかけ上CO2の出入が0の時の光量 [光合成量 = 呼吸量]日補償点: 日中蓄積する同化産物が夜に使われる有機物量と等しくなったときの日中の光量の平均値
吸収量 = 放出量 Rule: 補償点 < 日補償点 < 最小受光量 (最小受光量 × 1/10 ≈ 補償点)最少受光量: 植物が生存できる最少光量。一般に補償点の約1/10倍の光量
簡便法: 裸光 = 1 → 対象種がその何分の1の光量まで緑葉をつけるか
陽葉 = 小型、クチクラ層、柵状組織発達
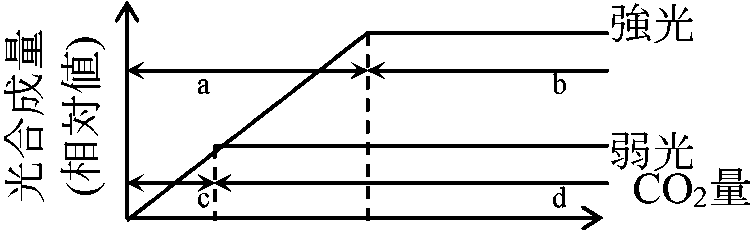
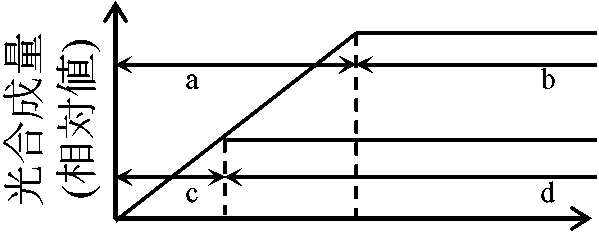
陰葉 = 薄く大形葉、柵状組織少 光合成支配要因ブラックマンの限定要因説 Blackman's law of limiting factors: 光合成反応速度は関係要因中もっとも不足した要因(=限定要因)に決定される要因: 光(波長), 温度, CO2量 (空気中の0.03%, 陸上で問題となることはまずない), H2O量(原形質の約3/4) 1) CO2量: 温度一定 – 最適温度
|
2) 光量: CO2十分
[光強度支配段階 = 明反応]と[温度支配段階 = 暗反応]の存在を予測 窒素同化と光合成 nitrogen assimilation and photosynthesis[窒素循環] 葉身窒素含量(N)と個葉光合成(P) ⇒ P ∝ NT ∝ Narea (Narea: 葉面積あたり窒素量 T: 葉の厚さ) Def. 比葉面積(比面積) specific leaf area, SLA = A/WL (m2/g)A: (葉の)葉面積 WL: 葉乾燥重量 Def. 葉面積当り葉重 leaf mass area, LMA = 1/SLA = WL/A≡ 比葉重 specific leaf weight, SLW (ほぼ死語) 葉面積比 leaf area ratio, LAR = A/W
W: 個体乾燥重量 (g) A: 個体葉面積 (m2) ∴ LAR = (WL/W)·(A/WL) = LWR × SLA = LWR/LMA SLAとLMAの意味構造指標: 葉厚、機械的支持組織量個体受光効率指標: 弱光 ⇒ SLA↓ / LAR↑ 葉寿命 ∝ SLA Def. 葉含水率 leaf water content , LWC = 葉水分重/葉総重量 水がなければ光合成はできない - 乾燥ストレス C/N比 (C/N ratio)陸上高等植物: 5-100 → 窒素利用効率指標タンパク質含む葉↓ 木質化した茎や根↑ 草本植物 < 木本植物 ⇒ 土壌のC/N比 |
C3植物 (C3plant), Rubisco系光呼吸する暗反応(カルビン回路)段階が最初C3化合物(PGA) = C3サイクル: 中間体のC3が重要なためこう呼ぶ [ 経路 pathway ] 食用植物はC3植物多 → O2分圧低下、CO2増加、RuBP oxydase阻害で光呼吸を抑え収量をあげる工夫をしても成功していない 葉形態: クランツ型 kranz (花輪): 維管束鞘細胞 bundle sheath cell 未発達か発達しても葉緑体なし C4植物 (C4 plant), Pep系1954, 1965 Kortschak et al.: サトウキビカルビン回路とは異なる光合成産物生成 1966, 1967 Hatch & Slack: イネ科・カヤツリグサ科・一部双子葉植物サトウキビ同様産物を検出 ⇒ C4ジカルボン酸回路 ≡ C4植物 光呼吸関与経路ない明反応初期物質がC4化合物でカルビン回路(暗反応)に入る= 光呼吸初期物質なく、直接O2使い酸化行う ↔ C3植物はリブロース2リン酸(C3)に結合しカルビン回路に入る ⇒ PEP + *CO2 → OAA → カルビン回路という道筋を使用 Ex. トウモロコシ・サトウキビ
CO2──┬──[エノールピルビン酸]┐
↓ │
[オキザロ酢酸] C4 [ピルビン酸]
↓ ↕
[リブロース2リン酸] [PGA]
┊ ↓
└┅┅┅┅┅┬────[グリセリン酸アルデヒドリン酸]
[グルコース]
12NADPH2 + 6CO2 → C6H12O2 + 6H2O + 12NADP____________ ATP↗↘ADP FpH2 + O2 → Fp + H2O2のような反応系を用いる 暗呼吸は電子受容体として働きエネルギーを生み出すが、光呼吸は非生産的であり機能的にも別経路 殆ど光飽和に達せず陽地では効率よい物質生産をする 葉形態: 非クランツ型: 維管束鞘が発達し、多くは葉緑体を持つ 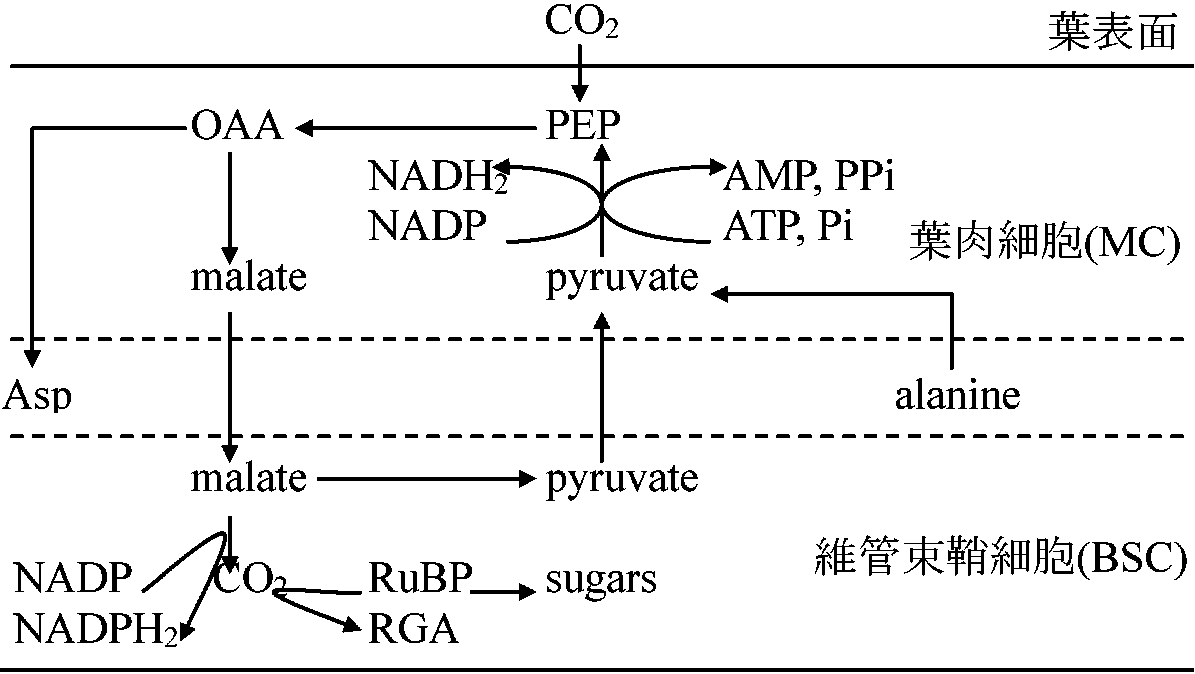
図. C4植物のCO2固定(例) C4型炭酸固定植物
第I群 NADP-malic enzyme型(脱水素酵素型) C4植物 vs C3植物C4植物 = 熱帯性植物 (Ex. トウモロコシ) vs C3植物 = 温帯性植物の大半サトウキビで最初のC4炭酸固定調査
単子葉植物 カヤツリグサ(35既知種), イネ(750) Panicum, Eleocharis, Atriplex: 属内C3/C4植物分化 = 炭酸固定は多系進化
C4固定に関係 = [PEP carboxylase, malic enzyme, pyruvate kinase] 葉肉mesophyllにある
(C3) RuBP carboxylase (C4) PEP carboxylase |
C3植物/C4植物の比較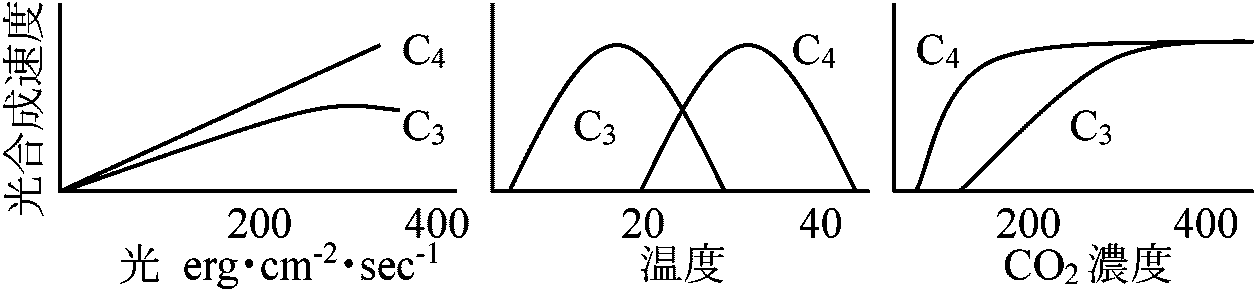
__________________
C3____________
C4_______________ Panicum milioides: 葉構造 = C4的 ↔ 機能 = C3的 → δ13C = -25‰ Hordeum distichon (温帯性植物): 葉C4型 ↔ 未熟果皮(pericarp) C3型Mollugo nudicaulis (ツルナ科/ザクロソウ科) 若葉(1-4枚目) C3型 ⇔ 古葉(11-13枚目) C4型 Eleocharis vivipara: 陸生状態 C4, 水生状態 C3CAM植物 (CAM plant)1974 Laetch WM: C4植物 = 雨期短い、熱帯サバンナ気候に適乾燥・高温が長期 = CAM植物の方が適 Def. 多肉植物 succulents: 水分貯蔵器官を持つ植物の総称
葉茎根いずれかが肥厚 - 水分貯蔵組織(柔組織)発達 科全てではなく多肉質植物の一部 → 多系統並行進化 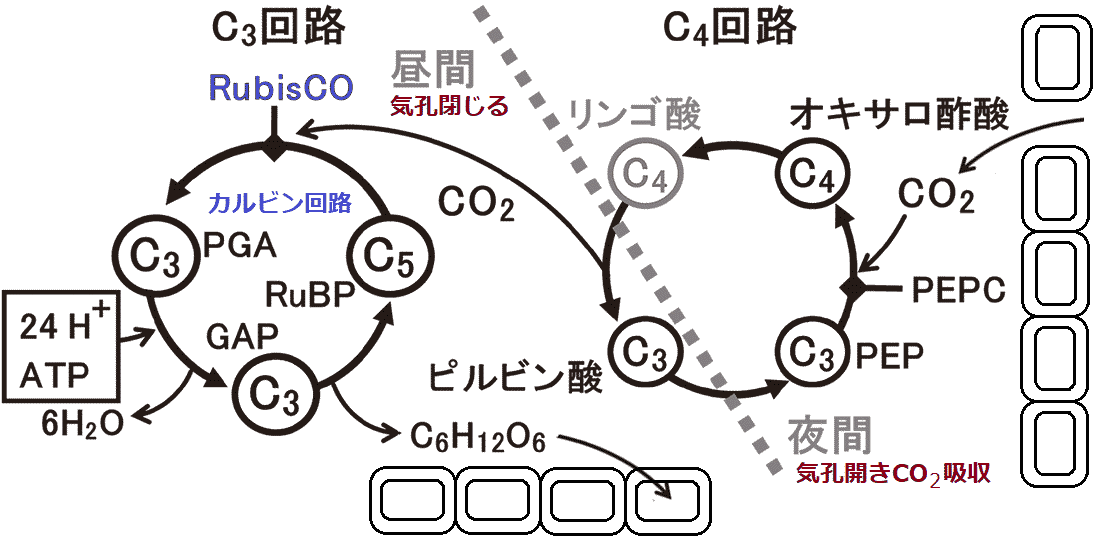 夜間に有機酸(リンゴ酸)貯え昼消費 → CAM存在 ⇒ C4回路を夜間、C3回路を昼間に行う Ex. 荒地草本: 光十分 ↔ 水分不十分CO2吸収に気孔開くと蒸発 ⇒ CO2吸収したい + 水分蒸発させたくない Ex. 午前中は光合成できるが、昼間から夕方に水不足で光合成行えない⇒ ジレンマ解消: CAM植物 = 気温低下し水分蒸発少ない夜間に気孔開きCO2吸収貯蔵 + 昼間は気孔開かず光合成 CO2は有機酸(リンゴ酸等)の形で貯蔵し光合成時にCO2に戻す CAM植物の特徴多様な光合成系を持つ - C3型とCAM型の相互互換が行われやすいEx. パイナップル: 水十分 = C3 ⇔ 乾燥 = CAM 1978 Osmond
可変性CAM植物 (facultative or inductive CAM plant) 進化高CO2濃度・低O2濃度時代: C3植物 ∵ CO2濃縮不要 →低CO2・高O2: C4植物分化 (C4 = CO2濃縮経路) + CAM (多系進化) 糖新生/グルコース新生 glyconeogenesis乳酸lactate、ピルビン酸pyruvate、クエン酸回路中間体、糖生成性アミノ酸等から、オキサロ酢酸oxalo acetateを経てグルコースを合成する過程
根本的に、糖新生系は解糖系の逆反応 |
|
強光(可視光照射) → 光合成反応阻害 = 光合成速度低下 (Powles 1984) 強光照射時にPSII電子伝達反応妨害され(条件でPSI)、行き場を失った光エネルギーによりタンパク質を損傷させる有害物質でき反応中心タンパク質D1の分解起こる ⇒ 光阻害の主な標的はPSII → PSII機能しなくなると光合成反応全体に支障 A. 光阻害機構1) 還元型光阻害強光で過剰電子伝達のため電子逆流し発生PS II複合体とシトクロムb/f 複合体間の電子伝達を受け持つPQは膜内に蓄えられ、PS II複合体から電子を受け取った後に膜内に戻る。過剰電子伝達が起こると電子を受け取っていないPQが膜内に不足しPS II複合体からそれ以上電子流れず、電子は逆流し戻って来るが、この電子は高エネルギーを持ち、タンパク質に有害な一重項酸素ができD1タンパク質は分解される 2) 酸化型光阻害PSII複合体が水を分解するが酸素を発生できない状態の時に光照射されると起こる。PSII反応中心では、集光性Chlに集められた光エネルギーはP680 Chlに渡される。P680は光エネルギーを受け取ると電子放出しP680+になる。P680+は強酸化力を持つが、通常は水から引き抜かれた電子を受け取り無害なP680に戻る(水を酸化し電子を引き抜く反応をしているのはP680+)。ところが、PSII複合体が水から電子を引き抜けない状態だとP680+が居座り周りの分子を酸化し、ついにはD1タンパク質が分解されるB. 光阻害防御1) 葉内で葉緑体の形を変化 + 位置移動2) 過剰な光エネルギーを別経路に流す
光(蛍光放散) + 熱(キサントフィルサイクル) + 物質(光呼吸) キサントフィルがエポキシ化やデポキシ化によって3つの形(ビオラキサンチン、アンテラキサンチン、ゼアキサンチン)に変化するサイクル |
非循環型電子伝達 (non-cyclic electron transport)1. H2OからPSII, PSIを経てNADP+への電子の流れ2. 水-水サイクル (water-to-water cycle) = 浅田回路 (Asada cycle) PSIIの量子収率調節のためのΔpH形成/暗順応条件において炭素同化系が失活している状態に光が照射された際の調節メカニズム 光過剰/炭素同化系停止 → 葉緑体でNADPHがCO2固定必要量以上生成 → PSIから過剰電子供給 → O2がNADP+の代わりに電子受容体となりスーパーオキシド(O2-)に還元 → … → H2Oに還元 1) 2H2O → 4e- + 4H+ O2PSIIのH2O光酸化によるO2発生、4e-が生じる 2) 2O2 + 2e- → 2O2- (律速段階)PSIによるO2還元とO2-生成 3) 2O2- + 2H+ → H2O2 + O2スーパーオキシドジスムターゼ(SOD)によるO2-の不均一化反応 4) H2O2 + AsA (or AsA) → 2H2O + 2MDA (or DHA)APXの触媒するH2O2のH2Oへの還元 5) 2MDA (or DHA) + 2e- + 2H+ → 2AsA (or AsA)PSIからの電子による酸化型AsAのAsAへの還元 MDA: モノデヒドロアスコルビン酸, DHA: デヒドロアスコルビン酸 (1)-(5)の合計: 2H2O + O2 → O2 + H2O
水-水サイクル: H2O由来電子により酸素分子が還元されH2Oを生ずる |
葉緑素抽出 (chlorophyll extraction)植物サンプル野外では採取したら速やかにドライアイスを入れたクーラーボックスに保管実験室に持ち帰ったら凍結乾燥しディープフリーザ(-60°C以下)に保管 凍結乾燥できるまでディープフリーザに保管 抽出試薬: ジメチルホルムアミド, DMF 手順
(Porra et al. 1989: 換算式は他にもあるので必要に応じ調べる) 計算式μM計算
Chl a = 13.43 × A663.8 - 3.47 × A646.8
Chl a = 12.00 × A663.8 - 3.11 × A646.8 パルス変調蛍光クロロフィル蛍光と吸収による光合成測定パルス変調蛍光測定 (PAM)= Pulse amplitude modulation学生実習 (student experiment)1. ヨウ素法2. 気泡計算法 3. エンゲルマンの細菌法
好気性細菌が酸素濃度の高い所に集まる性質(走化性)を利用し調べる 蒸散1. 実際測定2. コバルト紙法: 乾 = 青, 湿 = 赤 |
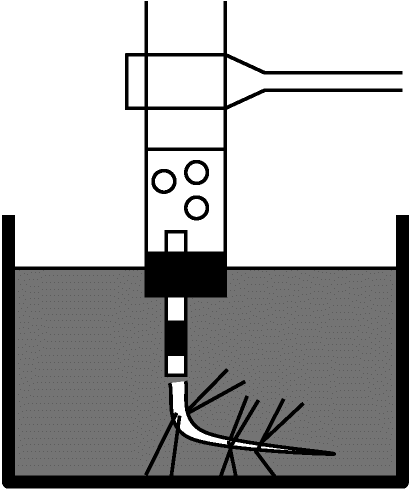
気泡計算による光合成測定内的・外的条件を一定に保ち、1つを変化させ変数間関係を知る。CO2濃度、温度を一定に保ち、光強度のみを変化させ植物のO2発生量を調べ光強度と光合成との関係を求める。簡単だが光合成機構理解の基本的実験である材料: オオカナダモ Elodea canadensis
原理: 水生植物茎切断時に出る気泡 ≈ 光合成によるO2 → 気泡発生速度 ∝ 光合成速度 → 気泡発生速度から光合成速度測定可能 内的・外的条件を一定に保つことに注意
同化度 実験結果: 温度23°C、CO2濃度M/100 KHCO3、照度3000-24000 lux
照度 (lux) | 測定値 | 平均 | 逆数 | 同化度 実験結果: 温度28°C、CO2濃度M/100 KHCO3、照度3000-24000 lux
照度 (lux) | 測定値 | 平均 | 逆数 | 同化度
|