(2025年6月21日更新) [ 日本語 | English ]
HOME > 講義・実習・演習一覧 / 研究概要 > 小辞典 > 生物学 > 微生物学
微生物 microorganism微小で肉眼で観察できない(ような)生物に対する便宜的総称 (< 100 μm)
= 観察に顕微鏡が必要な生物の総称 主な微生物群 = (微小)菌類 + (微小)藻類 + 原生動物 + 細菌 + ウィルス エンドファイト (内生菌, endophyte): 植物体内に共生する微生物
植物の病害抵抗性、環境ストレス耐性に関与 細菌 procaryotae, mychota, monera, anucleobionta原核細胞 prokariotic cellだが細胞内膜系としてメソゾーム mesosome、色素胞(クロマトフォア)chromatophoreの見られるものもある従属栄養的 heterotrophic → 腐生/寄生 (独立栄養 autotrophicなものもある) 光合成 photosynthesis か化学合成 chemosynthesis を行う 生殖(増殖) reproduction
遺伝形質組換 changes of gene types有性生殖はないが遺伝形質の組み換えがみられる
〇≡≡≡△
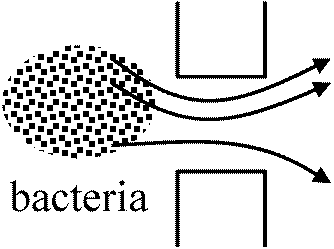
利用 use of bacteria for human
1675 Leeuwenhoek, Antonie van (1632-1723): 手製顕微鏡(倍率150倍) 糞便中から細菌・原虫発見 - 微生物発見(世界初) 1749 Ncedham: 肉汁に微生物自然発生1776 Spallmzani: 密封加熱実験で自然発生否定 1796 Jemer: 種痘の人体実験 1821 Fries: カビ分頼体系化 1826 Schwann et al.: アルコール発酵は酵母による 1807 Schwann: 発酵・腐敗の微生物要因説 1850 Mitscherlich: ジャガイモ褐変要因は細菌 1850 Davaine: 炭疽病家畜血液内に炭疽菌観察 1853 De Bary: 穀類のカビは寄生カビによる 1858 Pasteur: 乳酸発酵要因は細菌 (☛ 自然発生説否定) 1864 Lister: 消毒外科手術の開発 1870 Lewis: アメーバ赤痢病原体発見 |
1875 Brefeld: カビ純粋培養 1875 Bruce: ナガナ病患著からトリパノゾーマ発見 1876 Tyndall: 空気中に細菌確認 1876 Koch R (1843-1910): 炭疽病病原菌分離, 再発症実験 後に細菌部類学の基礎築く 1879 Neisser: 淋菌発見1880 Laveran: マラリア接合子を人血中に発見 1881 Pasteur, Roux: 炭疽菌による免疫実験 1882 Eberth-Gaffky: チフス患者よりチフス菌分離 1882 Schloesing Muentz: 土壌の硝化は細菌による 1882 Koch: ゼラチン平板培養法(平板法 plate method)開発
微生物培養(= 実験)可能となる (ゼラチンは後に寒天) 1888 パスツール研究所設立 1884 Gram HC (1853-1938): 細菌の差別染色法(グラム染色) 1892 伝染病研究所設立 1885 Hellriegel, Willfarth: 根粒のN2固定は細菌による 1888 Beijerinck: 根粒菌純粋分離. 集積培養考案 1889 北里柴三郎: 破傷風から嫌気性病原菌分離 1897 Buchner: 無細胞系アルコール発酵 1890 VonBehring, 北里: ジフテリア抗毒素発見 1890 Winogradsky: 独立栄養硝化細菌分離 1892 Iwanowski: タバコモザイク症からウイルス分離, 再感染実験 1895 Beijerinck: 独立栄養硫酸還元菌分離 1897 Frosch Loeffler: 動物から濾過性ウイルス発見 1898 Nocard Roux: 牛肺疾からPPLOを分離培養 1914 北里研究所設立(第1次世界大戦) 1898 Ross: ハマダラ蚊によるマラリアの媒介発見 1909 Orla-Jensen: 細菌分類に生化学性状を導入 1910 Ehrlich, 秦: 梅毒治療薬サルバルサン発見 1926 Kluyver et al.: 発酵に量子伝達概念導入 1917 D'Herelle: バクテリオファージ発見 1927 Griffith: 肺炎菌の形質転換発見 1929 Fleming, Chain A (1881-1955, 英): 青カビからペニシリン分離 1931 Van Niel: 紅色細菌の嫌気的光合成発見 1935 Domagk G Trefouel (1895-1964, 独): スルファニルアミド発見 1936 Stanley: タバコモザイクウイルス結晶化 1940 Fleming: ペニシリン治療実験成功 1940 Beadle, Tatum: アカバソカピの突然変異 1943 Delbrueck-Luria: 細菌の突然変異 1944 Waksman, Selman Abraham (1988-1973): ストレプトマイシン発見 土壌中放線菌から抽出 1945 Reyniers JA (米): 無菌動物飼育装置開発 - 微生物実験発展1947 Tatum, Lederberg: 大腸菌(E. coli)に性を発見 1950 Hungate, Robert E (米): 嫌気性細菌培養法(ロールチューブ法)開発 本方法によりルーメン内セルロース分解菌分離 1958 Meselson, Stahl: E. coli DNAの半保存的複製機作証明1977 Woese, Carl: 古細菌発見 1995 Venter Craig: 細菌全ゲノム配列決定 |
1. 夾膜と粘液層 capsules and slime layer原核生物細胞壁外側にある高分子有機物だが必須形態ではない。突然変異等で欠失することもあり機能未詳
大きさは200-500 nm程度が普通。夾膜-粘液層の境は不明瞭な種もある [ 繊毛 ] 2. 鞭毛 flagellum多くの微生物が鞭毛を有し、分類上重要形態 = 運動性 mobile配置: 極性 幾本かの蛋白質繊維が縄状にからまった構造。Eukaryotes’s compound flagellumの様な9+2構造を持ったものではない 構造: 3構成部 = 基部 + フック + フィラメント 鞭毛の集まり assembly of filament - flagellin subunit: どのように細胞壁に表れるかは不明 実験1. 細菌鞭毛 → [熱処理] or [高温処理] → 鞭毛再構成 実験2. A型鞭毛細菌(直毛) + B型鞭毛細菌(曲毛) → 鞭毛再構成 = A型実験3. A型鞭毛細菌の抗体を加える
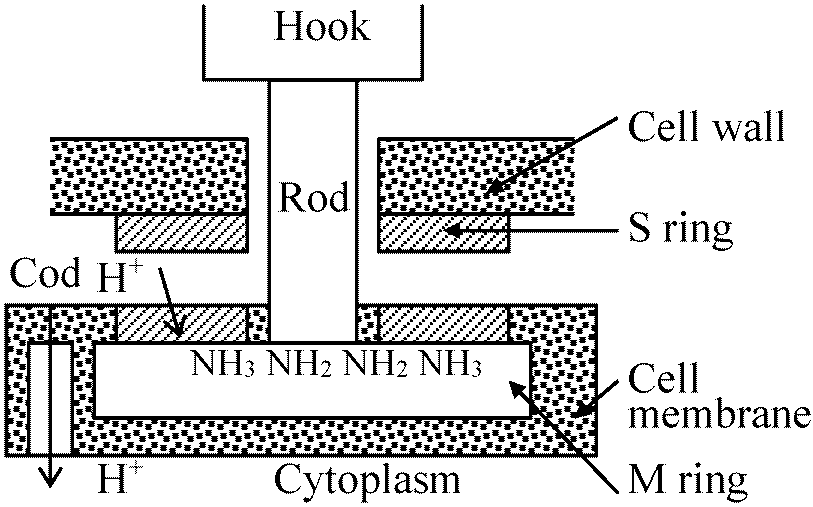
フックはCCW回転 = 細菌はフックの回転運動によって動く
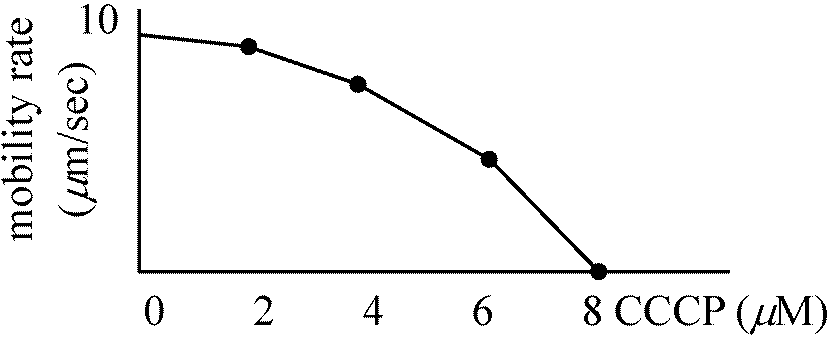
Ex. Vibrio cholerae: 80 μm/sec or 288 μm/sec
protein motive force, electrochemical potential – 膜内外の電位差とプロトン勾配に基づく
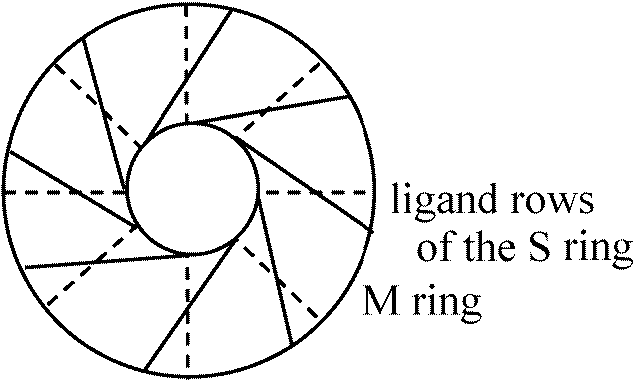
細胞内外のpH差(イオン濃度差)により細菌は運動する 化学要因グラム陰性菌: 下部リング(S, M)のみ = 上部対はロッド支持には不要 → SとMは必須器官ではない
走化性 chemotaxis正常移動は反時計回り counter-clock-wise (CCW)であるがtumblingが起こると時計回りclock-wiseになる誘引物質 attractantsg – CCW 忌避物質 repellents – 同軸回転tumbling増える
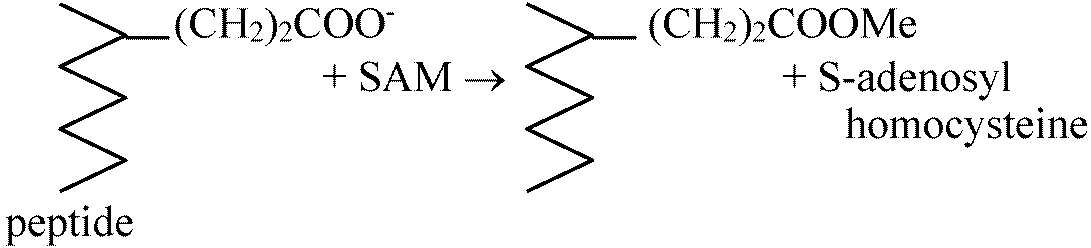
ATPあるいはMetが欠如すると細菌はchemotaxisを起こさない。さらにS-adenocylmethionine (SAM)を加えるとchemotaxisが起こる Fig. Methylated (-dimethylated)
各々の突然変異体は異なるMCP反応性を示す
MCP I欠如 (tsr mutants): セリン、グリシン、ロイシンに反応しない。温度変化の影響を受けない 3. 繊毛 pilus, pl. piliAnderson et al. (1949): 電子顕微鏡で発見。長さ30-250Å, 13-18 kDグラム陰性菌による研究が多かった Ex. Bordetella pertussis, Neisseria gomerrhoae グラム陽性菌: 一般に細胞壁厚く染色不適3分類群 Corynebacterium spp., Streptococcus spp.(連鎖菌), Mycobacterium smegmatisのみが知られる 分類 (classification)a) 細菌性線毛 bacterial fimbria or common pili幅4-10 nm, 長さ0.5-数μm, 100-300 pili/cell, MW = 13-18 kD Hydrohobic L-amino acids (Ex. PAK = 43.9%, F = 53%, Type I = 43.6%) 赤血球、白血球、イーストで繊毛はよく知られる 感染 infection: 微生物が体内に侵入し足がかりを確立すること。人間に感染する場合、腸管壁に付着し増殖を始めることが第一条件となることが多い Ex. Salmonella b) 性繊毛 sex pili (or fimbriae): 最初はE. coliで発見雄株 male strain: RNA phage(20面体)がpiliの回りに、棹状のDNA phageが先につく 雌株 female strain2種類確認 (共にplasmid性) → common piliと異なり数少なく1-2本/細胞程度
F-like pili (sex factor, or F factor): 大。R factorや幾つかのcolicin factorを有さぬものはpili形成なし 4. 核 nucleus (in prokaryotes)染色体 chromosome = バクテリア遺伝子MW 28 × 109 KDt, ca 3.2 × 106 塩基対 base pair (3.2 Mpb) 理論上3000種のタンパク質合成 = 3000遺伝子 (1970 Taylor : 310遺伝子マッピング → 317遺伝子)5. 細胞(内)顆粒 cytoplasmic granulesa) リボゾームr ibosomeb) 異染(ボルチン)顆粒volutin granule ポリリン酸 トルイジンブルーで染めると紫色にメタクロマチンmetachromatinが染まる cf. 細胞内顆粒中、リン酸が多量につながったもの: リポ核酸タンパク質、脂質、Mg++ Klebsieila, Azotobacter, Escherichia, Micrococcus, Nitrosomonas等が含む。細菌以外にChlorella, Euglenaでも存在
volutin granuleはエネルギー貯蔵(ATP補充)の役割なく、単なるリン酸のプールと考えるべき c) クロマトフォア(色素胞) chromatophore 細胞膜起源。光合成細菌特有。膜系持つ。60 nm |
d) Poly-β-hydroxylutyrate granule (PHB) Sudan blackで良く染色される。大きさ700 nm程度 細菌に加え、藍藻・原生動物の一部でも確認される PHBは貯蔵物質であろうと考えられている β-hydroxybutrateが600-2500分子つながる。膜で包まれているらしい Acetyl Co-Aがβ-hydroxybutyrateの起源 e) タンパク質結晶 B. thuringiensis胞子が斑点を作ることから確認。巨大結晶(機能不明)。タンパク質抽出 → リシンに対し特性 f) 液胞(ガス気胞) vacuole: Holobacterium (好塩菌)、他数種類で確認 g) 硫黄 sulfur 光合成細菌 H2S + CO2 → [CH2O] + S(蓄積) 化学合成細菌: 硫化化合物を酸化する
H2Sが欠乏するとSできない 6. サイトプラスム cytoplasm7. メソゾーム (mesosome)化学組成: 細胞膜との相違 – 量的差あるが質的差は殆んどない。リン脂質組成も質的差ない
タンパク質:_______________ 細胞膜 (50-57%) > メソゾーム
メソゾームの働きを呼吸中心と仮定するとmesosome/membrane ≫ 1.0にならないと辻褄が合わない UDP-Mar, Nac-pentopeptide 欠如, translocase欠如, polyglycerophosphate synthetase欠如 以上より仮説否定 単一の働きから複合した働きがあるのではないかと言われていた ⇒ 1970年代後半: アーティファクトであると結論 (experiment)8. 細胞外皮 (cell) envelopes= 細胞壁 + ペリプラズム(細胞周辺腔含む) + ペプチドグリカン + 細胞膜OCM細胞外皮 = これらの物質全体およびそれによって構成される構造 細胞の固い構造を保つため必要(細胞壁も構造保持機能持つ) ペプチドグリカンpeptidoglycanが構成の中心
細胞壁 構造a) グラム陽性菌の典型的構造 (☛ グラム染色)
━━━━━━━━━━ 200 Å = CW
▆▆▆▆▆▆▆▆▆▆▆▆▆▆▆ リン脂質でできている = CW 細胞外皮の構成因子 components of cell envelopea) ペプチドグリカン peptidoglycan網目状構造: GluNac (N. acetylgucosamine), MurNac (N. acetylmuramic acid) i) 骨格部: G-M-G-Mのβ-1, 4 bond ii) 側鎖部: D-Ala, L-Glu, L-Ala
-G-M-G-M-G-M-
NH2-Gly-Gly-Gly-Gly-Gly-Gly-COOH Ex. Erysipolothrix rhusiopathiae: NH2-Gly-Lys-NH2 ペプチドグリカンの厚さがグラム陽性菌と陰性菌で異なる
Negative: E. coli = 8 nm グラム陽性菌 + Butyrovibrio fibriosolvans (Gram陰性菌) lipo teichoic acid + teichoic acid グラム陽性菌全てがこの構造をとるわけではないが、陰性菌では1例を除きteichoic acidは未確認
Polytglycerol: (-OH残基1つ)
Cf. Glycerol
P-CH2 Piにより高分子化(約30単位)
Cf.
Polyribitol: (-OH残基3個) Glycerolと同じで30単位位まで高分子化する c) リポ多糖質 lipopolysaccharides:Outer-side (O-side) chain構造が細菌の特性を決定している
Ex. Salmonella LPS (outer core) 脂肪酸 CH3(CH2)nCOOH: C16, C18多く、更に不飽和脂肪酸持つもの多い 炭素鎖長 → 沸点高 (不飽和脂肪酸増加 → 沸点低) → 好熱菌・好冷菌膜構造はこれにより調節される 細胞外皮基本構造 basic structure of cell envelopesA) 膜モデル membrane modeli) Danielli model - 反証実験で殆ど否定 タンパク質粒子モデルも実測値と予測値の間に1:600の開きがあり殆ど否定 ii) シンガー・ニコルソンモデル Singer-Nicholson model B) サイトプラズミックメンブレン cytoplasmic membrane グラム陰性菌では内膜に等しい。膜系がないので細胞膜上で代謝がなされ、細胞膜は最も重要な系である C) 外膜 outer membrane グラム陰性菌のみ。酵素は昔はphospholipaseのみと言われていた → □ → □□ → □ + □ OMが肥大成長し、分裂面が出来るためには酵素が必要 Ex. E. coli: 3'-nucleotidase, 5'-nucleotidase, Mur-Nac-L-Ala amidase, N-acetylmuramidase等がOMに存在 これらは成長のための分解酵素を含む IM: 呼吸合成酵素群 → OMは生存上の重要性としてはIMほどではない 機能:
1. 構造を保つ
tetra lysine (MW = 530) – through ↔ penta lysine (MW = 660) – stop → MW 600前後で通過できなくなる
なぜ一定の場所に蓄積できるのか → そのための情報をpeptideが持つ?
I: 90% cytoplasm D) ペリプラスム periplasmic space, or periplasm 細胞壁と細胞膜の間の空間。この空間から特異的酵素見つかる → 抽出 高張液hypertonicにつける(osmic shock)するとleak out。次に低張液(水、希釈緩衝液等)につける 酵素: alkaline phosphatase (OM), cyclic phsophodiesterase (cAMPの5'-endを切断), 5'-nucleotidase タンパク質: 結合タンパク質(結合するアミノ酸決まっている) Ex. Leu, Ile, Val E) 粘着帯 adhesion zone 1963 Cota Robels, 1968 Bayer
Ex. 棹菌 rod-shaped bacteriaを10-20% sucrose溶液中に入れる → 原形質分離plasmolysis起こる
basal body of flagellum = motor on IM 3) ファージとの関係
ファージは粘着帯につく 元素主要構成元素 = C, N, P, S (珪藻 + Si)
一般に、微生物の方が大型生物よりもNとPに富む (CH2O)106(NH3)16(H3PO4) + 138O2 微量元素Fe: 微生物が必要な量は僅か(C:Fe ≈ 104)であるが制限要因となる 外洋では不足しがち (≈ 10-12 M) 高栄葉塩・低クロロフィル (high nutrient-low chlorophyll, HNLC) 土壌中には多量に存在 |
| グラム陽性菌 | |||
| 物質の種類 | 物質名(構成成分) | 菌名 | 夾膜の種類 |
| 多糖質 糖 アミノ酸 ウロン酸 アミノウロン酸 ベプチド、タンパク質 | dextran(グルコース) lavan(果糖) (アミノ酸) (糖、アミノ酸) (グルコース、マンノース、ウロン酸) (グルコース, N-acetylmannos-aminuronic acid) Hyaluronic acid (グルコサミン、グルクロン酸) r-D-グルタミルペプチド r-DL-グルタミルペプチド Mタンパク質 | Leuconostoc mesenteroides Streptococcus salivarius Bacillus megaterium Streptococcus pneumoiae B. circulans Micrococcus luteus Streptococcus spp. (A, C群) B. anthracis B. subtilis (natto) Streptococcus spp. (A群) | 細胞外粘質 細胞外粘質 夾膜 夾膜 夾膜 夾膜 夾膜 夾膜 夾膜 夾膜 |
| グラム陰性菌 | |||
| 多糖質 リポタンパク質、リポタンパク多糖質 | セルロース(グルコース) デキストリン(グルコース) (ウロン酸) Vi抗原(アミノウロン酸) (fucose, ガラクトース, hexuronic acid) Polyribophosphate | Acetobacter aceti Cluconobacter oxydans Klebsiella Salmonella typhi Escherichia coli Haemophilus influenzae | 細胞外粘質または繊維 夾膜 夾膜、ルーズな夾膜 ミクロ夾膜 夾膜 夾膜 ミクロ夾膜 |
[ 原核生物命名規約 ]
|
s.s. 原核生物 = 古細菌 archaea (archaebacteria) + 真性細菌eubacteria (一般に言う細菌 bacteria) s.l. 原核生物(s.s.) + ウイロイド viroids + ウイルス viruses 真核細胞から見た微生物真核細胞: 3箇所にDNA = 核 + ミトコンドリア + 葉緑体(植物)古細菌 archaebacteria: 高温・高塩・高イオウ等に棲む 太古地球環境に似る: 現細菌より古いという意味で命名(実は古くはない) → 始原菌 archaea と改名 = 第3の生物始原菌仮説 (Wosse et al.)真核生物 = 古細菌細胞内に真正細菌共生核遺伝子 ≈ 古細菌 → 細胞の元(説)
問題: 真核生物 = 核大、内部構造複雑
ミトコンドリア ≈ 酸素利用しエネルギー生産できる真正細菌(αプロテオバクテリア) スピロヘータの構造と真核細胞内の細胞骨格や中心体、精子尾等に構造上の共通性 分類法a) 形態分類(系統)グラム染色 Gram staining: 細胞外皮構造大きさ: 一般に長さ2-4 μm, 幅0.5-10 μm Thiosp sp > 50 μm, Haemoph influenzae 0.2-0.5 μm 鞭毛/細菌鞭毛 (bacterial) flagellum胞子染色 spore staining 寒天培養 agar culture shape, chromatogenesis, opacity, elevation, surface, edge観察 外部形態 form of bacteria (中間体はありうる)菌形態(顕微鏡レベル): 大きくは球菌と桿菌に分ける 球菌 coccus
単球菌: 細胞分裂後すぐに娘細胞は離脱 (以下は離脱せず集団形成)
短桿菌 |
bacterial ectoplasm: bacteriochlorophyll and carotenoid, etc.を有しphotosynthesisを行うbacteriaもおり、これらの成分はectoplasm中にある。この系列は独立栄養autotrophicである コロニー形態 (肉眼レベル) 形 form: circular, irregular, rhizoidと大きさsize 色素形成 chromogenesis Ex. S. marcensens = red-orange, S. aureus = cream-yellow, M. agilis = pink 透明度 opacity高さと盛り上がり方 elevation: flat, raised, convex 表面: smooth, rough, glistering, etc. 縁: entire, undulate, lobate, rhizoid, dentate 臭い odor b) 化学分類 chemotaxonomyO2必要性好気性 aerobic: 分子状態酸素存在下で正常成育 絶対好気性 obligate aerobic > 微好気性 micro-aerobic 嫌気性 anaerobic
偏嫌気性 strict anaerobic: 分子状酸素があると成育不可能 一般にチトクローム欠く - 分子状酸素を水素最終受容体としない 通性嫌気性 facultative anaerobic: 好気的/嫌気的条件いずれも成育 成長率ピグメント pigments 加水分解ゼラチン hydrolysis gelatin DNA, G/C content インビック試験 (IMViC test)代謝産物の検定により化学反応の存在を確かめ分類同定指標とする= インドール試験 + メチル赤試験 + ホーゲス・プロスカウエル試験 + クエン酸利用能試験 1. インドール反応 indole reaction2. メチルレッド反応 methyl red reaction 3. ホーゲス・プロスカウエル試験 Voges-Proekauer test 細菌がグルコースを酸化しacetyl carbonalを生産するか否かを観察 4. クエン酸利用能試験 citrate testCitrateを炭素源として用いているか否か c) 数量分類学 numerical taxonomy1957 Smith: 親和度Affinity (%) =(common characters)/(total characters, n > 50) × 100 1961-71 Cowan: 異端的記号分類法 heretical taxonomy
全種をコードで表わす |
[新タクソン, Bergey's Manual revised in 2001, ed. Garrity]
Kingdom Prokaryote 原核生物界Domain Bacteria 細菌 (Eubacteria 真正細菌)細胞球形 coccus - 棹状 bacillus細胞壁丈夫。有鞭毛では通常周鞭毛 グラム陰性-陽性(科でほぼ一定) 横分裂増殖(内生胞子形成するものあり) Phylum Aquificaeグラム陰性好熱性細菌温泉・海底火山 - 水素を酸化 Phylum Thermotogaeグラム陰性好熱性細菌熱水噴出孔・油田 - 有機物を発酵 Phylum Thermodesulfobacteria好熱菌、硫黄還元細菌Phylum Deinococcus-Thermus放射線耐性菌、好熱性細菌Phylum Chrysiogenetesグラム陰性Phylum Chloroflexi (緑色滑走細菌)Class ChloroflexiOrder ChloroflexalesChloroflexaceae (緑色糸状細菌): Heliothrix - 光合成行うOrder HerpetosiphonalesPhylum ThermomicrobiaPhylum NitrospiraPhylum DeferribacteresPhylum Cyanobacteria (藍藻細菌)Phylum ChlorobiClass ChlorobiaOrder Chlorobiales栄養体普通0.5-1.0 μmの微小単細胞細胞小器官未分化であり組織形成しない Chlorobiaceae (≈ 緑色硫黄細菌) 光合成細菌 = 酸素非発生型光合成 絶対嫌気性 Phylum Proteobacteria (プロテオバクテリア), syn. Pseudomonadota細菌最大の多様性リポ多糖(LPS)から成る外膜 - グラム陰性 好気性細菌(一部嫌気性) 水系菌種多 Class α-proteobacteria従属栄養海洋細菌真核細胞ミトコンドリアはこのグループの細菌由来(説) → 細胞内共生 Order RhodospirillalesOrder Rickettsiales (リケッチア)Anaplasmataceae: 多くは細胞内部でのみ生存可能= 細胞内共生菌 - 昆虫に高頻度 雄殺し male-killing, 単為生殖 parthenogenesis, 雄の雌化 feminization Order RhodobacteralesRhodobacteriiceae 紅色細菌細胞光合成色素有 - 主にグラム陰性好気性細菌 Order SphingomonadalesOrder CaulobacterialesOrder Rhizobiales (リゾビウム)(Hyphomicrobiales 出芽細菌, 統合廃止)Rhizobiaceae Rhizobium Frank 1889: 根粒菌 R. leguminasarum (エンドウ、ソラマメ) 酵母エキスマニトール寒天培地培養により2属に分ける
Rhizibium = 生育速く酸産生 R. japonicum (ダイズ) R. trifolii (シャジクソウ) R. lupini (Lupinus) R. meliloti アルファルファ菌 (ウマゴヤシ, シナガワハギ) Sinorhizobium Chen et al. 1988: 根粒菌(硝化細菌) Bradyrhizobiaceae Bradyrhizobium: 根粒菌 Afipia, Agromonas, Blastobacter, Bosea, Nitrobacter, Oligotropha, Rhodopseudomonas (カロチノイド含む)
Nitrobacter winogradskyi 硝酸菌: Cp. Nitrosomonas Phyllobacteriaceae Mergaert et Swings 2006Mesorhizobium Jarvis et al. 1997: 硝化細菌(系統解析Rhizobiumから独立) M. loti Jarvis et al. 1997: Nelumbo nuciferaと共生し根粒形成 Hyphomicrobicaceae 出芽細菌: 出芽 (or出芽 + 縦分裂) 細胞球形-卵形-楕円形-洋梨形。多くは集合体 Methylobacteriaceae (メチロバクテリウム) Beijerinckiaceae Methylocystaceae Rhodobiaceae Class β-proteobacteria (ベータプロテオバクテリア)好気性/通性細菌Order BurkholderialesAlcaligenaceaeAchromobacter (A. fisheri 発酵菌) Alcaligenes Bordetella (B. pertussis 百日咳菌) Taylorella Burkholderiaceae Comamonadaceae: Sphaerotilus (S. nataus = 水質汚染指標) Oxalobacteraceae: Janthinobacterium (土壌細菌、代謝能多様、低温等ストレス耐性高) Sutterellaceae 廃止: Chlamydobacteriales, Chlamydobacteriaceae 鞘細菌, 糸状菌(Ex. Sphaerotilus): 細胞は糸状体形成。一般に明瞭な鞘に包まれる。グラム陰性。極鞭毛持つ遊走細胞か不動性分生子生じる。内生胞子非形成 Order HydrogenophilalesOrder MethylophialesOrder NeisserialesNeisseriaceae: Neisseria (N. grorrhoeae 淋菌)Order NitrosomonadalesNitrosomonadaceaeNitrosomonas 亜硝酸菌 2NH3 + 3O2 → 2HNO2 + H2O + Energy (NH4OH → HNO2) SpirillaceaeGallionellaceae: Gallionolla (G. ferginea 鉄細菌 iron bacteria) 2FeCO3 + 3H2O + 1/2·O2 → 2Fe(OH)3 + 2CO2 Order RhodocyclalesClass γ-proteobacteriaグラム陰性病原菌多 Order Chromatiales (≈ 紅色硫黄細菌)ChromatiaceaeThiorhodaceae廃止 (Thio-: カロチノイド含む = 光合成行う) Ectothiorhodospiraceae Order ActithiobacillalesOrder XanthomonadalesXanthomonadaceaeOrder CardiobacterialesOrder ThiotrichalesThiotrichaceae: Beggiatoa (細胞単独性/糸状体。藍藻植物に似るものある(但しクロロフィルなし)Piscirickettsiaceae Francisellaceae Order LegionellalesOrder MethylococcalesOrder OceanspirillalesOrder Pseudomonadales シュードモナス旧 Class Schizomycetes 分裂菌(分生菌)綱含むPseudomonadaceae シュードモナス 色素生産あっても光合成色素持たない オキシダーゼ陽性 グルコース代謝能 胞子産生性 + 運動性 Pseudomonas (syn. Chlorobacterium 緑色細菌, 廃止) > 218種 多種有機化合物分解能 (分解者) → 炭素循環/バイオレメディエーション Azotobacter (A. chroocaccum 窒素固定)Azomonas Winogradsky (窒素固定) Order AlteromonadalesOrder VibrionalesVibrionaceae: Vibrio (Ex. Cholera group) 弧状-螺旋状の棹状 (V. möller, V. comma コレラ菌, V. parahaemelyticus 腸炎ビブリオOrder AeromonadalesOrder Enterobacteriales (腸内細菌)Enterobacteriaceae (腸内細菌): Enterobacter, Escherichia coli 大腸菌, Shigella 赤痢菌, Klebsiella (K. pneumoniae 肺炎棹菌), Salmonella (S. typhosa チフス菌), Yersinia, YokenellaE. coli (Migula) Cast. et Chalm.: グラム陰性桿菌、通性嫌気性菌
名前の通り腸内細菌 → モデル生物 Order PasteurellalesClass δ-proteobacteria, nom. nud. (δ-プロテオバクテリア)Order Myxobacteriales 粘液細菌栄養細胞薄膜。鞭毛なし。分泌粘液物質上滑走。グラム陰性括れ分裂。大部分は栄養細胞が集合し子実体fruiting body形成 栄養体は休眠細胞resting cell, microcyst, myxosporeとなり、子実体粘液中に埋もれたりマクロシストmacrocystという嚢中に形成される Order BdellovibrionalesBdellovibrionaceaeBdellovibrio Stolp and Starr (ブデロビブリオ): 偏性好気性細菌。汚水・土壌 他のグラム陰性菌内部に侵入し細胞質を捕食し増殖 Class ζ-proteobacteriaPhylum Firmicutes (グラム陽性細菌門)= グラム陽性低GC含量細菌Class Bacilli バシラスOrder Bacillales バシラス芽胞形成偏性好気性(一部通性嫌気性)グラム陽性桿菌 Bacillaceae Bacillus (B. subtilis 枯草菌, B. anthracis 炭疽菌) – 細胞は真直ぐな棹状 Planococcaceae Caryophanaceae Caryopahales 廃止 Caryophanon (C. latum – 細胞円柱状-円板状。グラム陽性。ゴニジア) Listeriaceae Staphylococcaceae: Staphylococcus, Staphylococci (S. aureus 黄色ブドウ球菌), Gemella, Macrococcus, Salinicoccus Sporolactobacillaceae Paenibacillaceae Alicyclobacillaceae Thermoactinomycetaceae Order Lactobacillales (ラクトバシルス)Lactobacillaceae ラクトバシルスLactobacillus: L. apodemi (タンナーゼ産生), Paralactobacillus, Pediococcus Aerococcaceae Carnobacteriaceae Enterococcaceae エンテロコッカス Enterococcus: 腸球菌・乳酸菌 Leuconostocaceaae |
Streptococcaceae (連鎖球菌) Streptococcus: S. gallolyticus(タンナーゼ産生),S. lactis 乳酸発酵菌(連鎖球菌), S. mutans 虫歯菌, S. pyogenes 化膿連鎖球菌, S. pneumoniae, Syn. Diplococcus pneumoniae 肺炎球菌 Lactococcus incertae sedis: Acetoanaerobium, Oscillospira, Syntrophococcus Class Clostridia クロストリジウムOrder Clostridiales クロストリジウム主要腸内細菌(全種ではない)Clostridiaceae Clostridium Prazmowski 1880: C. tenani 破傷風菌, C. botulinum ボツリヌス菌 Order ThermoanaerobacterialesOrder HalanaerobialesOrder NatranaerobialesOrder Thermoanaerobacteriales多系統群Class Erysipelotrichi エリュシペロトリクスOrder ErysipelotrichalesClass Negativicutes ネガティウィクテス従属栄養性、偏性嫌気性、グラム陰性Order Acidaminococcales アキダミノコックスOrder Selenomonadales セレノモナスOrder Veillonellales ベイロネラVeillonellaceae Rogosa 1971Veillonella parvola: ヒト、ラット、ウサギの口腔・腸管 Class Thermolithobacteria Sokolova et al. 2007 テルモリトバクテルOrder ThermolithobacteralesThermolithobacteraceae: Thermolithobacter (鉄還元する偏性嫌気性菌)Class Limnochordia Watanabe et al. 2015 リムノコルダClass Tissierellia ティッセレッラPhylum Tenericutes テネリクテスClass Mollicutes モリクテス細胞壁欠く = 微小 + 動植物寄生Order Mycoplasmatales マイコプラズマ (Classとして分類)一般の細菌が持つ細胞壁欠く (ウイルスも欠く) - 特異的無細胞培地で増殖 → コロニー非常に小 ≈ 250 μm-1 mm 肉眼識別困難。培地に血清や腹水を加えるのは多種でステロールsterol合成能力がないため 多様。運動性なし。グラム陰性 生殖不明 (基本的に2分裂増殖 - 大きさ、形まちまち) Mycoplasmataceae マイコプラズマ Mycoplasma, incertae sedis: M. pneumoniae 原発性異型肺炎病原体 Erysipelotrichaceae: Erysipelothrix AL, Holdemania Order Entomoplasmatales エントモプラズマOrder Acholeplasmatales アコレプラズマAcholeplasmataceaeCa. Phytoplasma Firrao (ファイトプラスマ) Order Anaeroplasmatales アナエロプラズマOrder Haloplasmatales ハロプラズマPhylum Actinobacteria (アクチノバクテリア. 旧, 放線菌)グラム陽性。DNA中GC含量高い菌種含む(= 高GCグラム陽性菌)Class ActinobacteriaSubclass ActinobacteridaeOrder Actinomycetales (放線菌/放射菌)従来: 菌糸形成に特徴付けられる分類群 = 系統的位置: 細菌と糸状菌の間
栄養体多少とも分枝した菌糸体生じるが菌糸幅 < 1.5 μm Suborder ActinomycineaeActinomycetaceaeSuborder CorynebacterineaeCorynebacteriaceaeDietziaceae Gordoniaceae Mycobacteriaceae: Mycobacterium (M. tuberculosum 結核菌, M. leplae ライ病菌) Nocardiaceae Tsukamurellaceae Williamsiaceae Subclass RubrobacteridaeOrder RubrobacterialesSuborder RubrobacterineaeRubrobacteriaceaeSubclass CoriobacteridaeOrder CoriobacterialesSuborder CoriobacterineaeCoriobacteriaceaeSubclass SphaerobacteridaeOrder SphaerobacteralesSuborder SphaerobacterineaeSphaerobacteraceaeSuborder Frankineae放線菌群から独立Frankiaceae フランキア Frankia Brunchorst: ☛ 放線菌根性植物 (nifD-K: フランキア特異的) 1888 Brunchorst: 放線菌根性植物の根粒菌 = Frankia (真菌類とした) 1902 Shibata (柴田): ハンノキ、ヤマモモ根粒組織の細胞記載 = 放線菌 1932 Krebber: Alnus根粒 = 放線菌 (成功は否定的) 1978 Callaham et al.: フランキア菌分離に初成功 Comptonia peregrina (L.) Coult.根粒から分離 + 接種により根粒形成 形態(顕微鏡レベル): 繊維状栄養菌糸、胞子塊、小胞体繊維状栄養菌糸 vegetative hyphae (Lechevalier & Lechavalier 1990)
0.5-2.0 μm w、分枝稀、気中菌糸形成しない。菌糸色様々 胞子: 1-5 μm、運動性なし、耐熱性なし 小胞体 vesicle: ほぼ円形の有柄構造物窒素固定酵素(ニトロゲナーゼ)存在部位 1987 Baker: 交互接種試験 - 菌株を多種植物に接種し根粒形成評価1) ハンノキ/ヤマモモ、2) モクマオウ/ヤマモモ、3) グミ/ヤマモモ、4) グミ 1996 Swensen, 1999 Jeong et al.: rbcL解析複数祖先種由来(≠ 一祖先種) 2007 Benson & Dawson: 根粒形成にnif解析を併せ3群に分類
1) Betula, Myrica, Casuarina Microsphaeraceae Sporichthyaceae Acidothermaceae Kineosporiaceae Suborder Micrococcineaeグラム陽性土壌細菌Micrococcaceae Bogoriellaceae Rarobacteraceae Sanguibacteraceae Brevibacteriaceae Cellulomonadaceae Dermabacteraceae Dermatophilaceae Dermacoccaceae Intrasporangiaceae Jonesiaceae Microbacteriaceae Beutenbergiaceae Promicromonosporaceae Suborder MicromonosporineaeMicromonosporaceaeSuborder PropionibacterineaePropinibacteriaceaeNocardioidaceae Suborder PseudonocardineaePseudonocardiaceaeActinosynnemataceae Suborder StreptomycinaeStreptomycetaceaeSuborder StreptosporangineaeStreptosporangiaceaeNocardiopsaceae Thermomonosporaceae Order Bifidobacteriales (ビフィドバクテリウム)BifidobacteriaceaeBifidobacterium ビフィズス菌(俗) B. longum BB536 (特定保健用食品, 森永) Phylum PlanctomycetesClass ChlamydiaeOrder ChlamydialesChlamydiaceaeParachlamydiaceae Simkaniaceae Waddliaceae Phylum SpirochaetesClass Spirochaetes細胞細長く5-500 μm, 柔軟、屈曲性あり最低1回螺旋を巻くグラム陰性菌。横分裂 Order Spirochaetales 螺旋菌(スピロヘータ)旧科名 Treponemataceae廃止Spirochaetaceae: Borrelia (B. recurrentis 回帰熱ボレリア), Treponema (T. pallida 梅毒菌) Serpulinaceae Leptospiraceae: Leptospira (L. icterohaemorrhagiae 黄疽出血性レプトスピラ) Phylum FibrocacteresPhylum Acidobacteria (アシドバクテリウム)培養困難: 2綱11属17種 (2009)
環境中16S-rRNAから推定される多様性は大 Phylum Bacteroidetes (バクテロイデス)グラム陰性菌腸内細菌叢主要構成菌(水系、土壌等にも分布) Class BacteroidesOrder BacteroidalesBacteroidaceaeRikenellaceae Porphyromonadaceae Prevotellaceae Class Flavobacteria好気性桿菌 (2–5 μm × 0.3–0.5 μm)GC含量 = 32-37% Order FlavobacterialesFlavobacteriaceaeClass SphingobacteriaOrder SphingobacterialesSphingobacteriaceaeFlexibacteraceae Phylum Fusobacteria (フソバクテリウム)Class FusobacteriaOrder FusobacterialesFusobacteriaceaePhylum VerrucomicrobiaPhylum Dictyoglomi |
|
単細胞生物: 個体形態的変化(突然変異除けば)なし = 未分化の連続的変化 = 個体群 populationとして扱うのが普通 B. subtilisは内生胞子1つであり、クローンにより増やしたコロニーを調べると遺伝的制御が確認される。コロニーは螺旋を描きながら増えるので、その機構が調べられている 増殖速度測定法1) 物質量変化 a) 乾燥重量、総N量、タンパク質N量、b) 濁度、c) 容積2) 細菌数(1より直接的) a) 生菌数 viable counting b) 総菌数 counting total cell number (total count)
顕微鏡下で一定区画内の菌数を数える。実際には死菌をも数えるため過大評価となる ii) membrane filter使用 貧栄養湖等では菌数極少 → 濾過濃縮して検鏡 iii) 粒子測定器coulter counter使用 – 細菌数測定以外にも利用
膜間に電圧をかけておき、そこを細菌が通過すると微少だが抵抗が増す。その電位差ΔV(通過した細菌数に比例)の度合いから粒子数(菌数)測定
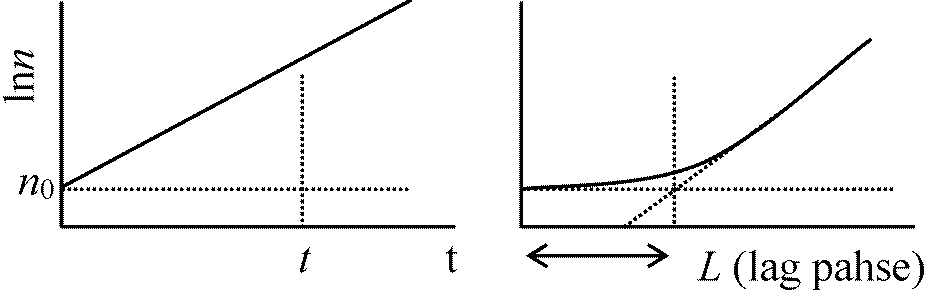
孔に平行に細菌は通過したと過程(これと実際の通過経路とのずれが大きいほど誤差は大) → aはAより十分小さいと仮定 = v/V·(ρ – ρ0)/ρ ≈ v/V [一般成長式] 成長曲線 growth curveN: 細菌数/物質量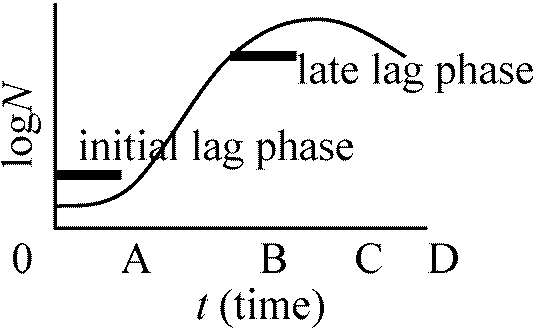 0-A: 誘導期 lag phase
0-A: 誘導期 lag phaseA-B: 対数期 log phase(増加直線) or 指数期 exponential phase B-C: 定常期 stationary phase: 一般にはこの相で終わる C-D: 減衰期 decline phase (= 死滅期 logalithmic death phase) Lag phaseの存在- なぜ存在するのか細菌は濃くするとlag phase短い → 前のコロニーに何か原因があるらしい 定常期の存在は栄養の枯渇が第一原因 + 他に代謝産物の蓄積等 最大個体群成長サイズは種により決まっているが、この原因は不明 増殖速度 growth rate細菌分裂時間は条件が揃えば大体決まっている = 平均世代時間 T最初n0個体存在 → t時間後n個体となる n = n0 × 2t/T → logn – logn0 = t/T·log2 ∴ (logn – logn0)/t = log2/T 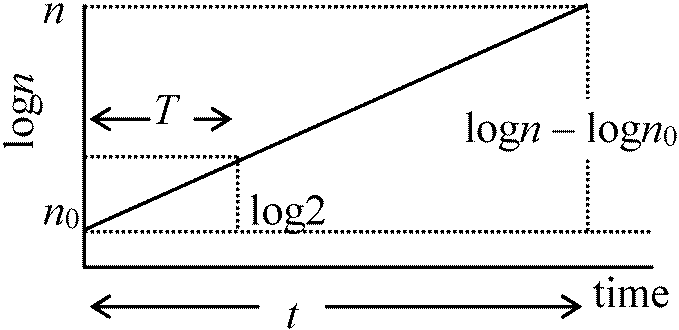
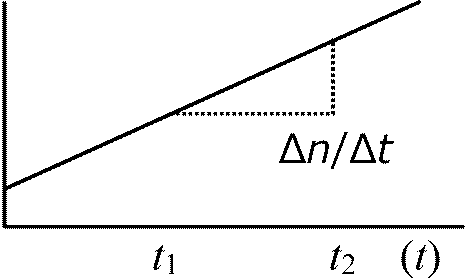
Δn/(logn2 – logn1)/(t2 – t1) = log2/T
このようにして平均世代時間Tは求められる 比増殖速度 specific growth ratedn/dt = kn (k: 比増殖速度 constant = 1/n·dn/dt)n = n0ekt ln(n/n0) = kt → lnnをもとに対数プロット → 傾き = k/2.303
前式からlog2/T = k/2.303 ∴ T = 0.693/k 一般に下記のように表わす
細菌数 n: k = 1/n·dn/dt (logn – logn0)/(t – L) = log2/T (T, L: 変数) → lag phaseでの測定よりT, Lは求まる 1949 Monod: log2nを縦軸とする = 2倍時間 double timen代目はlog2n0 + nで表わせる(最初は0代) 1955 Finey: 2を底とする対数表作る (結局無駄)最大生産量 maximum population or yield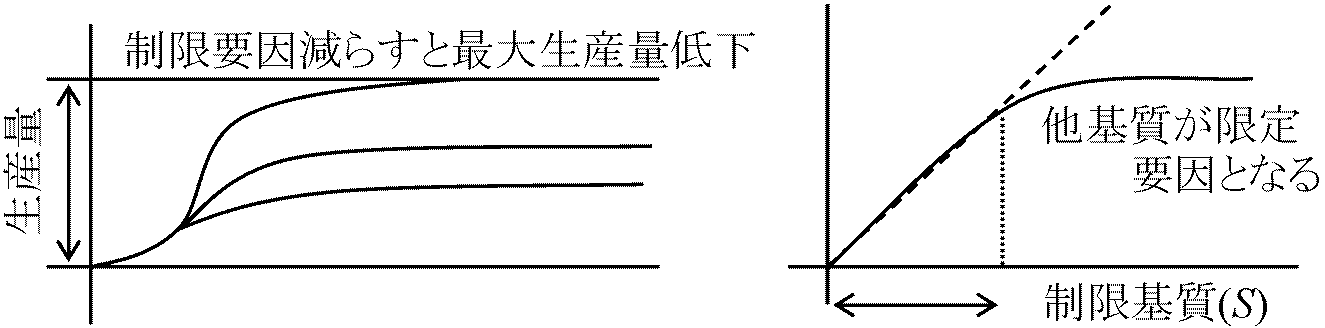 逆に最大生産量から制限基質濃度分かる(生物検定, 生物試験法 bioassay) vitamins, amino acids, purines, pyrimidines, etc.で広く生物検定は利用 (生成した細菌量)/(消費された制限基質量) = Y Y: 収量 yield (収量定数, yield coefficient or constant) モル増殖収量 molar growth yield: 1モルの制限基質で増殖できる菌数= 生成ATP数 [反論: 定量的関係はない] 共代謝 co-metabolism (共酸化)エネルギーを取出さずに物質分解・構造変化行う(= 同化異化に無関与)
生成物質: 他微生物が利用可能な状態へ変化 ⇒ 物質循環系形成
バッチ系: 容器中に培養液を入れ微生物を閉鎖的に増殖 連続培養 continuous culture細菌をある定常状態に保っておく培養方法利点: 齢の揃ったものを連続的に集められる Ex. バッチ法 batch method
μ = (1/x)·(dx/dt) 基質依存 → cf. Michaelis constant: v = V·S/(KM + S) |
KS: saturation constant 集積培養 enrichment culture: 自然環境かその採取試料の培養
発酵型微生物 zymogenic group: 外部栄養物により増殖 選択培養 selective culture: 培地栄養条件調整し特定微生物を増殖 Ex. 無機塩培養 - 化学合成無機栄養微生物発見 増殖と基質濃度dx/dt = -Y·(ds/dt) … (2)Y = -dx/ds (Y: yield constant) D = f/V (h-1) D: 希釈率 dilution rate, f: 流速 (ml/t), V: 培養液の濃度(m/V) 1/D (hr): 存在時間となるもし細菌が増殖しないと洗い出される: 洗い出しの速度 –dx/dt = Dx (槽内の菌数) … (3) 増殖していると:dx/dt (槽内の変化) = μx (増殖) – Dx (洗い出し) … (4)
状態_定常状態_______菌数増加__菌数減少(洗い出し) ds/dt = DsR – Ds – μx/Y … (5) (1), (4)より dx/dt = x[μm·s/(Ks + s) – D] … (6) (5), (6)より ds/dt = D(SR – s) – μm/Y·s(Ks – s) … (7) 定常状態の時: dx/dt = 0, ds/dt = 0 ∴ μm·s/(Ks + s) = D D(SR – s) = μmx/Y·s/(Ks + s), Y(SR – s) = x, s = Ks·D/(μm – D) → μm, Ks, Yが分かればx(菌数), sが分かる x~ = Y(SR – s) = Y{SR – Ks·D/(μm – D)}, s~ = Ks·D/(μm – D), W~ = x~ 洗い出し: DW~ = Dx~ Km = μm, Sk = ks 同調培養 synchronous culturet: 平均世代時間mean generation time = T → 個体群: 様々なステージ
Seitz filter (Cu製, Ptメッキ) Upper layer: Toyo filter #1, 2 layered, Lower layer: Toyo filter #126, 18 layers, etc. E. coli, lag phase 1 l → final 2 ml (10'' cells)濃縮, 50 mlの培地を2.5 ml/secで洗い出す
Small-sized bacteria__________Large-sized bacteria ショ糖密度勾配法 SDG method (Mitchison & Vincent 1965)
10-40% 500 g, 10 min → large cells, 4% of total bacteria Ex. Millipore filter (type GS)
菌が引っかかる____rinse: 分裂後の若い細胞を集める
ii) 光 illumination: 光強度変える iii) 飢餓 starvation: 貧栄養 → 富栄養 同調細菌より酵素(ATC, asparate transcarbamylase)抽出 → 高活性 → 酵素生成も同調 貧栄養微生物 oligotrophs: 貧栄養条件下で増殖可能
1: 富栄養-貧栄養で増殖可 同調培地での増殖(成長)固形培地で成長測定は困難だが、コロニーカウント及び純粋培養によりコロニー形態観察には便利成長の化学的(栄養的)条件
化学栄養細菌類: 無機塩類、CO2、(光) 1) 培地組成 medium composition合成培地 synthetic medium: 組成明らか完全培地 complete medium 自然培地 natural medium: 組成不明物含 Ex. molt extract, meat extract, peptones, potato extract, milk, yeast extract 栄養培地 nutrient medium (選択培地 effective medium)Haemophilus sp. (= blood必要): for H. influenzae ()内がbloodで必要
___________________X factor_______V factor 2) 無機塩類 mineral salts細菌の構造を保つ3) 炭素源 carbon source最も普通な炭素源はD-glucose
利用できない細菌も結構いる Ex. Vibrio ABE-1, Flavobacterium 4) 窒素源 nitrate sourceアミノ酸合成に必須 (☛ 窒素循環)5) 発育系 growth factor (≈ bacterial vitamin)生理的活性必要有機物 = 生理的活性が可能となる最低限があればよい。主に補酵素co-enzyme(アミノ酸等で自身で合成可能なもの、構成要素として存在するものは発育系と言わない) Inositol: fungi, yeast, Actinomyces israelii (細菌ではこの種のみ) Choline (恐らく膜形成に関与): Pneumococii, Mycoplasma (inositolも必要) Oleic acid (C18:1): Clostridium diphtheriae Vitamin 自然条件での増殖自然(自生)培養 natural culture: 自然条件下で増殖させたもの ↔ 純粋培養→ 自然培養(微生物生態学)と純粋培養(微生物生理学)との乖離 1936 Rossi / 1930 Cholodny: 埋没スライド法 buried slide technique
スライドグラスを土壌や水中に埋没 → 顕微鏡観察 毛細管を埋没させ毛細管中に微生物を収める |
微生物叢/細菌叢 microbiotaマイクロバイオーム microbiome: ある範囲での微生物全体⇒ 微小生態系 microcosm: 微生物含む微小生態系 Ex. 水溜、金魚鉢 水と湿度微生物は、常に水を介しO2, CO2等の気体を摂取 (乾燥土壌でも微生物は水の膜を介し気体摂取)腐敗や発酵は、基質に含まれる水が関係 - 微生物生育に水分不可欠 基質中水分は、形態から結合水、自由水に分類
結合水: 基質構成成分であるタンパク質や炭水化物と固く結合した水 自由水の割合 = 水分活性 water activity (aw) = 蒸気圧 vapor pressureを測ればよい(Scott 1953) 水分活性: aw = P/P0 → 溶液蒸気圧(P)と純粋蒸気圧(P0)の比
概ね基質平衡相対湿度(ERH)の1/100 → aw = P/P0 = ERH/100
好乾性 xerophilic: 陸上植物、サボテン、トノサマバッタ
浸透圧好塩菌水分の多い所 = 普通低張な所に住む Ex. グラム陽性菌 = 20 atm, グラム陰性菌 = 5-10 atm. 広塩性 euryhaline: 浸透圧変化に広い耐性↔ 狭塩性 stenohaline: 耐性範囲狭い 細菌表層は丈夫(例外: Mycoplasma – 細胞壁ない)Cf. 菌類菌糸 – burstしない: 内部浸透圧を下げburst避ける, プロトゾア– 積極的排水, 花粉発芽 水圧 water pressure1万m以下の深海で生存可能な細菌存在 → 100-160気圧で成長異常 = 一概に好圧性barophilicか不明
Ps. bathycetes, 1000気圧, 3°C → Lag phase = 4 months, generation time = 33 days, stationary phase = 1 yr?
Ex. 多価アミラーゼ: 6000 atm/cm2にすると変性する pH (pH)普通の細菌はpH 5-9.5程度が最適 (≈ 0という分類群もいる)好酸性細菌 acidophilic bacteria
Thiobacillus → H2SO4, Acetobacter → CH3COOH Bacillus alkalophillus (ca pH 11), B. alkaliphillus → 多量のNH3必要 |
1942 Gale EF
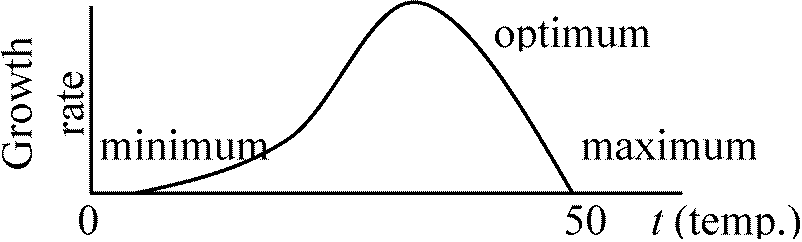
E. coli, amino acid dehydrogenase → 塩基性, Deaminase → 酸性 温度 temperature基本的温度 ordinal temperaturea) 熱致死温度 thermal death temperature
→ 液体培地(水溶液)中で10分おくと死ぬ時の温度
凍結防止剤 anti-freezerを加え凍らせず測定 成長に最適な温度 – 成長率曲線より求める d) 最大温 度maximum temperature: 測定自体は容易
最適温度: 理論上左右対称 菌根ヘルパー細菌 mychorrhiza helper bacteria, MHB1994 Garbaye: [定義] 菌根菌の菌糸生育と植物-菌根共生を促進する細菌2005 Schrey et al.: Streptomyces - 菌糸伸長
促進 -菌根菌(ベニテングタケ) ↔ クロマツ菌根圏: Bacillus, Ralstonia 微生物群集 microbial community広域: 水平分布(北半球) 赤道 → 北極 ≈ 垂直分布 低標高 → 高微生物密度↓ 耐乾性胞子形成細菌↓ 放線菌↓ 糸状菌↑ 大気: 飛散(上昇気流・ジェット気流等) ≠ 増殖 → 地表 ∝ 多様
細菌: 塵や埃に付着し大気中で生存か?
好圧性微生物 barophile: 数百気圧以上の圧力でのみ増殖可 - 深海 冬期 = 対流 → 鉛直方向種組成変化小 ↔ 夏期 = 滞留 → 大 1972 ZoBell: 海水中藍藻 = 300種 → 30種 浮遊生物 + 残り 付着生物 periphyton 土壌: 表層: 有機栄養微生物 - リター・腐植利用 (藻類・原生動物の殆ど)
深部: 化学合成無機栄養微生物
好気性細菌(O2消費) → 嫌気性細菌 (遷移) 植物根棲息種: 生きた植物内でのみ棲息可能 群集解析0. 成長測定応用
a) コロニー培養: 培養困難な細菌では不可 2. DGGE: バンドパターンのみ 3. 遺伝子解析 a) メタ16S/18S解析: 16S/18S rRNA - 遺伝子配列解析進む ⇒ 配列データベース 細菌等からゲノムDNA調製し16S rRNA該当領域の塩基配列解析 - データベース利用し分類群推定可能 利点: 安価、少量、系統組成欠点: PCRバイアス, 分解能(属まで), 系統機能不明 生物多様性 biodiversity ∝ アンプリコン配列 16S amplicons多様性 機能的冗長性 functional redundancy: metagenomeを介し測定
biodiversity ∝ function (or redundancy) 利点: 系統組成/遺伝子機能、バイアス稀 欠点: 参照配列依存、目的依存 |
分類群 (taxa): 遺伝子(タンパク質)は高等生物に近い - 真正細菌から独立
環境 (environments)1)温度好熱菌 (好熱性細菌 tempmophilic bacteria)多くは温泉、深海熱水孔付近で発見: 深海 = 高圧 → 100°C以上の水存在40-100°C: 好熱性細菌(普通は70deg;C程度で成長) → 超好熱菌: 100°Cで働く酵素を持つ Pyrolobus fumarii: 121°Cのオートクレーブ処理でも死なない
8-50°C: 中温(常温)菌 ≠ 始原菌 細胞抽出物cell free extractを煮ても変性しない 1. タンパク質(酵素含む)自身が熱に安定 - PCR (1993 Mullis ノーベル賞)
熱安定 thermolysin: 表面に2、内部に2のCa++の存在。Ca++を除くと熱失活する
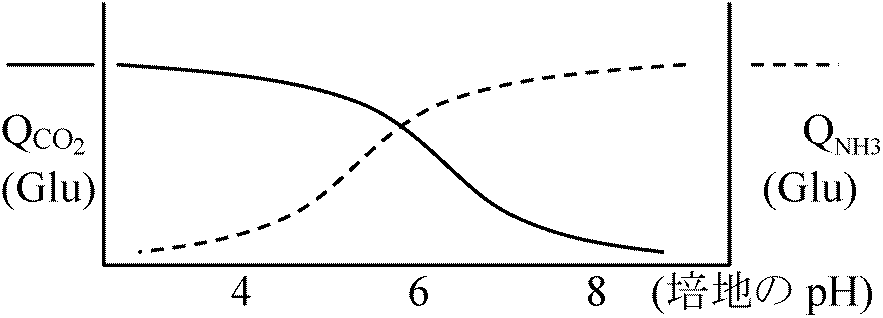
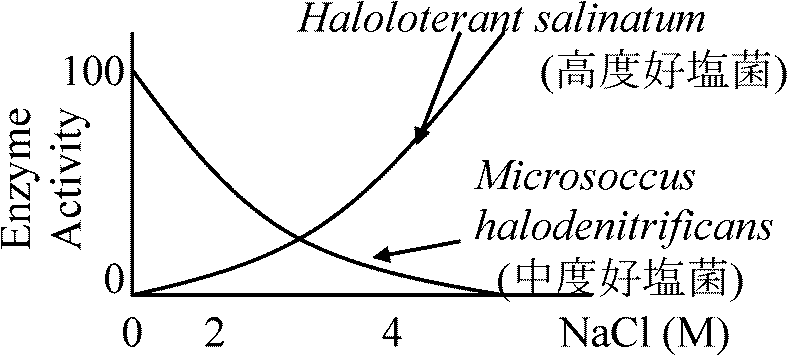
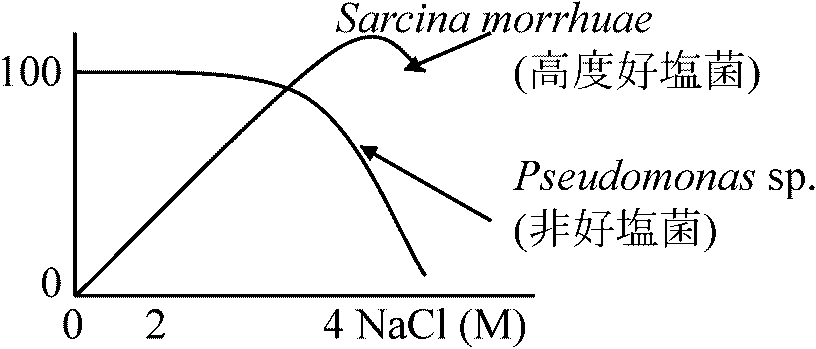
aldolase = optimum temperature 95°C (97°C 20' 100% active) 一部の細菌では脂肪酸が熱に安定な糖脂質に置き換わっている 3. 補修早い(証拠なし) ↔ 好冷菌: 殆ど不明2) pH3) 塩分弱好塩性生物: 1-6% NaCl ⇔ 中好塩性生物: 6-15% NaCl⇔ 超好塩性生物: > 15% NaCl 高度好塩菌 = 好塩性 (塩漬保存: 高濃度塩中で通常微生物増殖不可を利用)死海(イスラエル)、グレートソルト湖(米国)等の塩湖、天日塩・岩塩中から分離
低好塩性 ___ 2-5% NaCl (海水3.3-3.7%)
P = CRT = 4 × 0.082 × 293 = 96.104
Pseudomonas B16 (low halophilic bacteria): [Inner] Na+, Cl+ ≈ Outer solution, [Innter] K+ > Outer solution → K+が10-30倍に濃縮
NaCl (M) 外液 細胞内 比率(%) → Na+の関与: 物質取り込み、構造維持、酵素の働き(酵素調べれば分かる) Micrococcus halodemitrificans = 中好塩性細菌
Ex. 1. Succinate DH
低張度ではgNaで外へ捨てる。細菌では、外周近くが高濃度で中央に行くにつれ低濃度になる |
1978 Brown AD: 多数カルボキシル基を有する
K+と結合したもの活性あり → 非結合のが非活性
______→ -K+ → 非好塩性細菌タンパク質でも、コハク酸等を使いアミノ基をカルボキシル基と置換させると好塩性時に活性化タンパク質となることもある 好塩性細菌リボゾームタンパク質 halophilic bacterial ribosomal proteins, HBRP
酸性 = 塩必要な1要因
Halobacterium (好塩菌)
始原菌: 5S RNA → 酸性タンパク質多
バクテリオロドプシンにより光エネルギーを用いてプロトンを汲み出す → プロトン勾配形成 4) 圧力耐圧生物: 大気圧以上でも生存はするが増殖は見られない
_好圧生物: 中程度の圧力条件下(10-80 MPa)で増殖 メタン菌(-生成菌/-産生菌/-生成古細菌) methanogen湖沼・海洋底泥(ヘドロ)、湛水水田土壌、ルーメン、シロアリ後腸、嫌気発酵汚泥、温泉源・深海熱水噴出口等から分離 = 有機物最終分解者= 絶対嫌気性古細菌: 酸素存在環境で増殖しない (自然界メタン生成主要因) 実験培養困難: 特殊ガラス容器に培養液入れ、加圧し水素ガス注入閉栓 → 孵卵器内培養
人工増殖困難 → ドブ環境 = 好適環境(自然生態系壊すと復元困難)
化学反応(メタン生成)でエネルギー獲得 (※ 系統分類群ではない)
水素や蟻酸 → メタン: Methanobacterium
Ex. 球菌 + 桿菌。運動性細菌 + 非運動性細菌 メタノフラン (MFR), テトラヒドロメタノプテリンZ (H4MPT), 補酵素M (HS-CoM), 補酵素B (HS-CoB), 補酵素F420 (F420, Maxabs = 420 nm + 蛍光 → 顕微鏡下で他細菌との区別に有効), 補酵素F430 (F430) 中温性メタン生成細菌 = 増殖遅 →多くの生理生化学的研究は高温性メタン生成細菌で行われる 偏性嫌気性: 大気レベル酸素濃度暴露 - 死滅= 増殖遅 → 取扱い難しく生態的研究遅れる 嫌気リアクター発生メタンガス = エネルギー源 →他環境からの発生メタンガスは大気放出 = 温暖化 メタン菌による生活排水浄化生活排水 → 下水道管 → 下水処理場 → 雑物除去 → 微生物活性利用し有機性物質分解* → 放流*浄化 = 微生物処理: 微生物と空気の接触をよくし好気性微生物の勢い強める → 分解速度早まる 浄化 → [残渣 = 多くの沈殿物発生] → 通常、焼却し灰化か、醗酵させ肥料残渣を嫌気的環境にしメタン菌を増殖させるメタンガス変換施設 → 飲料可能水と僅かな残渣のみに変換可 メタン生成細菌 vs 水素生成細菌共通点: 嫌気性細菌 ↔ 相違点: 性質 - 特に利用する栄養分は異なるメタン菌 ↔ 水素生成菌: 炭水化物やタンパク質等高分子を好んで利用するが、酢酸等利用できない 水素生成細菌は、メタン菌と違い、セルロース等固形物を構成成分であるブドウ糖まで分解し、ブドウ糖を水素、CO2と酢酸に変換 水素は水素燃料電池で電気変換可 + CO2は光合成微生物でデンプン変換可 |
|
1858 de Bary HA: 動菌類 Mycetozoa 提唱 (∵ アメーバ運動、微生物捕食) 1869 Brefeld: Dictyostellium ⊂ 植物界(菌類 Eumycota, vetus definitio) 1969 Whittaker: 五界説 - 粘菌を菌界 fungi に含める 1998 Cavalier-Smith: 分子系統解析 - アメーボゾア門 Amoebozoa に属す Phylum Myxomycota (Myxobionta) 変形菌(粘菌)植物門生活史: 栄養生活相-生殖相明瞭
栄養生活相 = アメーバ相: 細胞壁なくアメーバ状
= 偽変形体pseudoplasmodium (アメーバ状に集合したもの) か変形体plasmodium形成 生活環の一時期にアメーバ相持つ + 別時期に肉眼で見えるキノコ様胞子嚢形成し胞子散布する 生殖: 被膜した胞子spore用いる - 耐性型細胞。包嚢cyst形成従属栄養 = 光合成能力なし → 真菌植物、原生動物に類縁関係求められる 食作用phagocytosis (エンドサイトーシスendocytosisの一種)による 変形菌和名: かつてはムラサキホコリカビ等、ホコリカビを最後につけた → 変形菌 ≠ 菌類 ⇒ カビを落とし、ムラサキホコリ、サビホコリ等と呼ぶ 系統 phylogeny63属400種程度
アラクシア型 ディクチオ型
細胞性粘菌 真性粘菌 細胞性粘菌
↑ ↑ ↑
寄生粘菌類 │ エキノステリウム アシトステリウム
↑ │ ↑ ↑
│ │ │ ┌----------┘
│ │ 原生粘菌類 ラビリンチュラ類
│ │ ↑ ↑
(植物的) ← 鞭毛プロチスタ → (動物的)
Class Acrasiomycetes アクラシスOrder Acrasiales アクラシス腐植土壌中、植物遺体中、糞中等栄養生活相: 単核アメーバ → 細菌や菌類を捕食し増殖
リマックス型: アメーバ細胞は1葉状仮足出し運動速度速く、細胞の前後の区別が明瞭 ☛ Dictyosteliaceae タマホコリ Dictyostelium Brefeld タマホコリ D. discoideum Brefeld キイロタマホコリ モデル生物: 細胞集合・分化、細胞運動・シグナル伝達 → 進化/創薬 Polysphondylium OliveClass Labyrinthulomycetes ラビリンチュラOrder Labyrinthulomycetes ラビリンチュラ(網状粘菌)有性生殖未詳栄養生活相: 単核アメーバの緩い網状群体。網状群体の一部にアメーバが集まり休眠胞子塊形成 |
Labyrinthulaceae Labyrinthula Class Myxomycetes 変形菌 (真正粘菌/真性粘菌)= 粘菌類(古くは動菌Mycetozoeaとも呼ぶ)栄養生活相: 複相核を持つ変形体plasmodiumと呼ぶ裸の原形質 生殖: plasmodiumより1-多数の子実体形成。ここに多数の被膜胞子を作る。胞子は発芽し単相・単核アメーバmyxoamoebaか遊走細胞生じ、接合し複相核形成し、その後分裂を伴わない核分裂で再び変形体となる Subclass Ceratiomycetidae ツノホコリ (原生粘菌)生活環の一時期に1個体で立ち上がり1-数個の胞子をつけた微細な子実体sporocarp形成Order Ceratiomixales ツノホコリ胞子: 子実体表面微(小)突起mucroに1個ずつ外生的に形成成熟胞子: 4個の単核相含み、胞子は発芽し4核持つ1個の原形質塊を生じ、有糸分裂後に8個の遊走細胞を生じる。減数分裂は胞子中で行なう Ceratiomyxaceae ツノホコリCeratiomyxa J. Schrot. 1889 ツノホコリ: C. fruticulosa (O. F. Müll.) T. Macbr. ツノホコリ Cavosteliaceae
Ceratiomyxella tahitiensis: 多核アメーバ - 1核を残し他の核は崩壊 → 3回核分裂し8核となる → 細胞質分裂で8個の鞭毛細胞形成 Subclass Myxogastromycetidae 真正変形菌 (真正粘菌)胞子は外皮peridiumに包まれた子実体中に内生的に形成子実体形状区分 (形状多様)
胞子嚢 sporangium Order LicealesOrder EchinostelialesOrder TrichialesOrder PhysaralesPhysarum モジホコリ: P. polycephalum Schwein 1822 (モジホコリ)Order StemonitalesSubclass Stemonitomycetidaeとする見解Stemonitaceae Stemonitis ムラサキホコリ Phylum Plasmodiophoromycota ネコブカビ維管束植物根に細胞内寄生 → stachelとrohrと呼ぶ特殊オルガネラが出現し宿主細胞内へ侵入宿主枯死後、子実体から胞子を土上に散布 雌雄の胞子zoosporeが融合しできた2倍体のplasmodiumが増殖 不等鞭毛2本 マスチゴネマmastigonemeなし Class Plasmodiophoromycetes ネコブカビOrder Plasmodiophorales ネコブカビ寄生粘菌 Ex. Plasmodiophora brassicae |
[protocol]
グラム染色 Gram stain(ing)細菌分類鍵である細胞外壁層を色素染色する方法(改良法多)= 陽性・陰性は構造(+ 生理機能)の違いを反映したもの 細菌分類基準グラム陽性 gram-positive: 紫色に染まるペプチドグリカン層厚 → 染まりやすい |
グラム陰性 gram-negative: 紫色に染まらない = 赤く見える
ペプチドグリカン層薄 + 外膜有する → 染まりにくい |
実習 (strain observation)材料
染色 staining (含, グラム染色)試薬
染色液
スライドガラス (slide or cover glass)に水を一滴垂らす → slantより菌を白金耳でとりslide glassにこすりつける(水滴が均一に薄く濁る程度に) → 風乾 固定: 風乾したものをバーナーの上の炎中を1, 2往復させる 単純染色 simple staining染色液乗せる(Methylene blue = 30-60 sec., Dilute fuchsin = 30 sec.) → 水洗: ビーカに水道水を入れ軽く濯ぐ(菌付着面の反対面に緩く水が当たるよう流水洗浄でも可) → 風乾 → 検鏡: カナダバルサム包埋 |
胞子染色 spore staining脱脂: 5% chromatic acidを乗せる(1-3 min) → 水洗 → 加温染色: carbal fuchsinを乗せ2-3 sec.バーナ上にかざす → 自然にさます → 水洗 → 脱色: 1-3% H2SO4を乗せる(2-5 sec) → 直ちに水洗 → 染色: methylene blueを乗せる(30-60 sec) → 水洗 → 風乾 → 検鏡総じて時間を計っておくと、後々便利 (西沢・菅原) 鞭毛染色鞭毛への硝酸銀付着が目的。鞭毛が水洗中取れ易く注意材料: Pseudomonus sp. 試薬
残り90 mlの硝酸銀を加える 手順: 1: 若いslantに滅菌水2-3 ml入れ1-2 hr放置. 2: 菌泳ぎ出し滅菌水薄く白濁したものをピペットで少量とりslide glassに乗せる. 3: 風乾(火焔滅菌と固定しない). 4: 媒染・固定: 第1液を乗せ2-3 min放置. 5: 静かに水洗. 6: 染色: 第2液を載せる(数秒). 7: 水洗. 8: 風乾. 9: 検鏡 水中菌観察理学部水道水(地下水)を無菌的に採取し菌数測定準備: 300 mlフラスコ、広口瓶、スプレッダー、寒天オートクレーブ 手順
|
|
脂質の細胞中の存在場所選択 エーテル・クロロフォルム(CF)・ベンゼン・エタノール・メチルアルコール・アセトン等で抽出
有機溶媒として一般にエーテル・エタノールの混合物を使用し、酵素変成を避けるために室温で抽出 脂質の種類により適当な溶媒に溶かし、カラムクロマトグラフィー・PC・TLC・GC等を用い分離・固定 細菌からの脂質抽出分析目的: 脂質抽出原理を理解し技術を獲得。定性分析を行い脂質の特徴理解材料: Pseudomonas aeruginosa (緑膿菌): 薄緑色蛍光を発する → 手順 (procedure) |
濃度 concentrationCF層をナス型フラスコに取りロータリーエバポレーターで濃縮 (< 25°C)乾固直前にロータリーエバポレーターを止め脂質を目盛付試験管に移す。ナス型フラスコにCFを適量(毎回同量)加え壁面を3回繰返し洗う 注) 事前に遠心管の重量を(cold room or ice bath等の)低温下で計っておく共栓は試験管にN2-gasと一所に封入。保存の際には-20°C 定量: 最終産物1 mlを秤量ビンに取りロータリーエバポレーターでchroloformをとばした後、2日間乾燥させdry weightを算出。今回サンプルは3つ用意。ブランクは1 ml NaCl dry weightを測定。別個にDDを測定しておく結果
B.W.W.-Blank Blank B.W.W.-Blank Lipid |
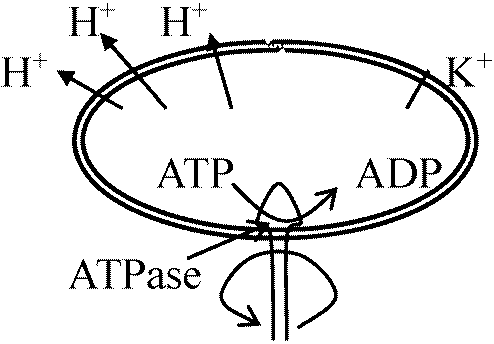 ATP合成時と同様、エネルギーによりプロトンが入って来る際に鞭毛が回る
ATP合成時と同様、エネルギーによりプロトンが入って来る際に鞭毛が回る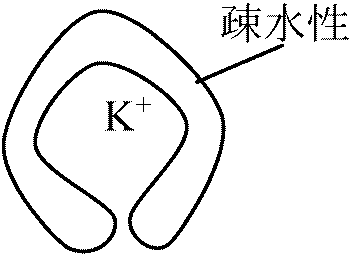 疎水的な膜でもK+を含むと膜透過性がある
疎水的な膜でもK+を含むと膜透過性がある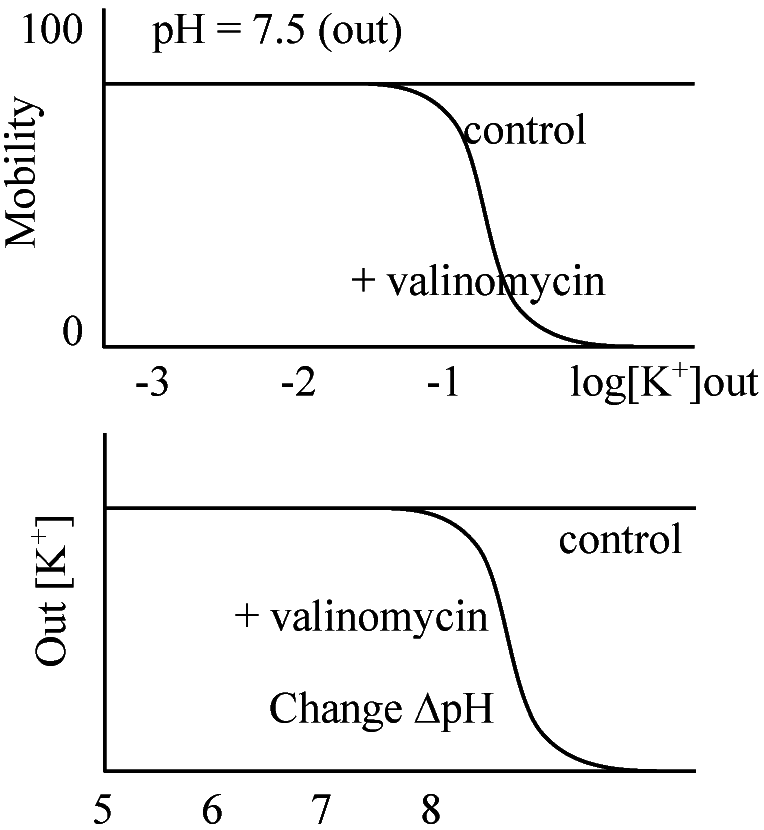
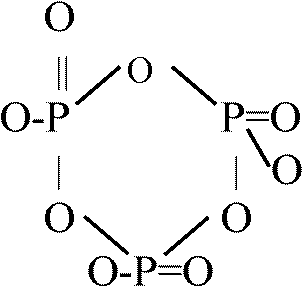 トリメタリン酸
トリメタリン酸
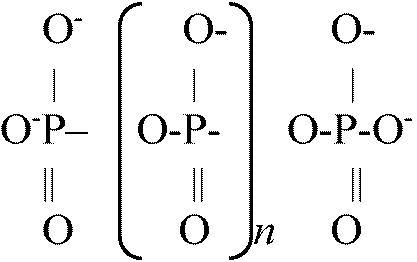
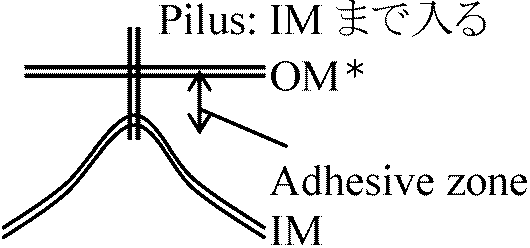 *: OM (phospholipids, liposaccharides, proteins – IM内で合成)
*: OM (phospholipids, liposaccharides, proteins – IM内で合成)