(2018年11月25日更新) [ 日本語 | English ]
HOME > 講義・実習・演習一覧 / 研究概要 > 小辞典 > 生物学 > 細胞
|
細胞の大きさ
卵黄 (Φ): ダチョウ 75 mm, ニワトリ 30 mm
植物: 100 μm l
分裂組織: 5-10 μm Φ バクテリア: 1-10 μm Φ Ex. タマネギ発芽根: 0.3 mm Φ, 3-4 mm l細胞 20 × 18 mm Φ |
|
|
真核生物 eukaryote = 真核細胞 (多細胞 + 単細胞)
ミトコンドリア内エネルギー生産 細胞内組織有さず真核生物のミトコンドリアmitochondria機能は細胞膜内側
細菌類、藍藻類 = monocellular
glucose, (NH4)2SO4, P, S, Mg → 1回分裂に50分毎
1) 大腸菌にinsulinを作らせる
原核細胞
真核細胞 |
細胞内膜系全てER起源。ER表面糖タンパク質組成 ⇒ ゴルジ体・リソゾーム等発生決定細胞基質 cytoplasmic matrix無構造。タンパク質・酵素・RNA含む。収縮性タンパク(アクチン・ミオシン)。原形質流動後形質 metaplasm原形質が変化(原形質の1種) = 原形質物質交代で生じた細胞内非活性物質+ それに基づく構造 Ex. 細胞液、澱粉粒・卵黄粒・脂肪粒・種々結晶体・色素・乳液 細胞器官器官に相当な働きを持つ細胞内構造鞭毛 (運動): ミドリムシ 繊毛 (運動): ゾウリムシ 眼点 (視覚, 光感受): ミドリムシ・ゾウリムシ 収縮胞 (体内余剰水分排出): ミドリムシ・ゾウリムシ 食胞 (消化器官的機能): ミドリムシ・ゾウリムシ・アメーバ 細胞含有物 Ex.貯蔵物質: デンプン粒・タンパク粒・脂肪粒・イヌリン・卵黄・グリコーゲン排出物質: シュウ酸石灰、炭酸石灰結晶体、乳液、アルカロイド、油滴、樹脂 浸透圧維持物質: 各種塩類 表. 細菌、古細菌、真核生物の定義に関わる諸特性
特性
細菌
古細菌
真核生物 SU: サブユニット。CP: クロラムフェニコール、SM: ステレプトマイシン、KM: カナマイシン |
細胞壁 cell wall, CW: 植物(細菌・藍藻)細胞特有な後形質
機能 function体制(構制): 個々生物体(系統)における構造の基本形式 - 相称や器官配置の主な様式を示す
Ex. 真正粘菌 → 不定形 amorpha: 体制とは関係ない 機械組織 mechanical tissue: 骨格として働く Ex. 膜組織、角組織 構造 structure中層にペクチン質を含む3層構造 → 内外層はセルロース主体で全透性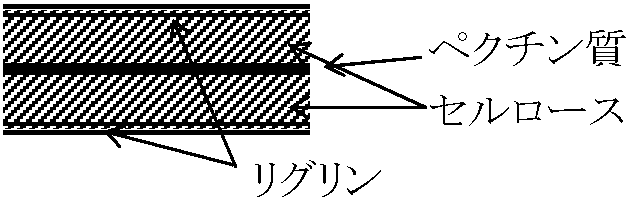 a) 中間層(中葉・中層) middle lamella (intercellular substance)
a) 中間層(中葉・中層) middle lamella (intercellular substance)
細胞間でどちらの細胞に属するとも言えない部分 →
セルロース, プロトペクチン, ヘミセルロースが主成分。植物全て共通
成分_______ヘミセルロース ペクチン物質 セルロース タンパク質 脂質 三層二次壁 three-layered secondary wall
仮導管 tracheid・導管 vessel: 3層二次壁から発達 → 柔組織細胞 parenchyma: 二次壁ない
リグニン = 木化
主成分: リグニンlignin → 木(質)化(肥大化) lignification = リグニン蓄積によりsecondary wallを形成 ヘキソース: D-グルコース(Glu), D-ガラクトース(Gal), D-マンノース(Man) ペントース: L-アラビノース(Ara), D-キシロース(Xyl) |
ウロン酸: D-ガラクトロン酸(GalA), D-グルクロン酸(GlcA), 4-o-メチル-L-グルクロン酸(Me-GlcA) メチルペントース: L-ラムノース(6-デオキシ-L-マンノース), L-フコース(6-デオシキ-L-ガラクトース) 二糖類(グルカン) glucan 1) UDP-Glucose → UDP + Sucrose: sucrose synthase (EC 2.4.1.13) 2) UDP-Glucose → UDP + Sucrose-6P: sucrose-P synthase (EC 2.4.1.14) Sucrose-6P + H2O → Sucrose + Pi: sucrose-phosphatase (EC 3.1.3.24) cf. Sucrose → Fructose + Glucose: invertaseセルロース cellulose ヘミセルロース hemicellulose デンプン starch アミロース amylose: α-1,4結合で300-1000個つながっている アミロペクチン amylopectin: Amyloseの鎖で所々にα-1,6結合の枝がある
α-1,6 α-1,6 α-1,6: 結合するときはQ酵素を用いる 酵素活性 enzymatic activitypectin methyl esterase-1,3-glucanase 1965 Lampont: この理論とタンパク質の存在は現在否定
セルロース中から特殊なアミノ酸を抽出 → 細胞壁の分化に関与すると考えた
_Arb___Gal Hydro Serine (Lysine, Xyrosin, Valine, Threione) 植物の細胞間連絡機構単細胞 = 当然、連絡機構なし → 群体 = 細胞間関係あり →多細胞(植物) = 密接な細胞間連絡機構必要 a. 原形質連絡 plasmodesma: 細胞壁を通し細胞同士を連絡する細い筋
一次壁のみの細胞間に普通 - 小胞体が連絡し合う (EM観察から) c. 師板 sieve plate・師域 sieve area d. 原形質膜(細胞膜)による連絡 |
|
動物細胞より植物細胞の方が顕著
動物: 電子顕微鏡でやっと観察できる 液胞成分を液胞外に取り出すと破壊される + 植物細胞では細胞壁が除去しにくかった → 液胞研究遅れた 液胞膜 vacuolar membrane or tonoplast: 植物細胞で特に発達し内部に細胞液を含む 細胞液 cell sap: 細胞膜に包まれた液体。液胞内に存在する溶液で(古くなった)植物細胞に特に発達 |
機能
+ 種子: 発芽タンパク質は特殊な液胞(protein body)に貯蔵 |
|
細胞構造を分化させているものが膜 → 単位膜(Unit membrane): 3層構造
この場合tonoplastとplasma membraneは同じもの 形質膜は液胞と細胞質とを区別する膜 - 液胞膜と構造的に似る
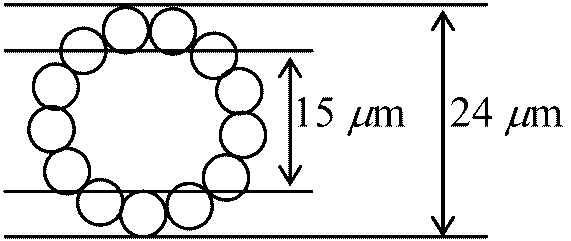
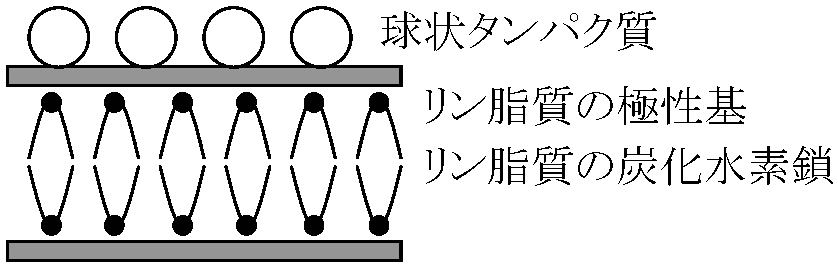
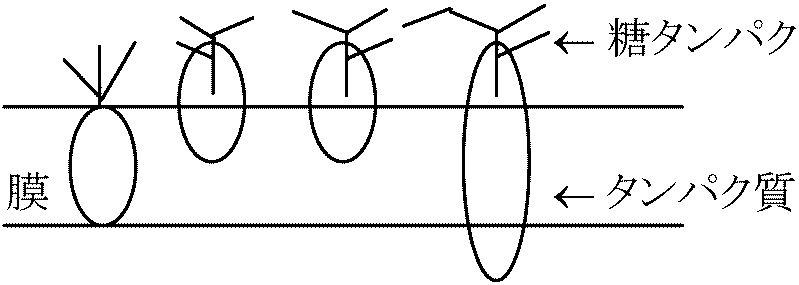
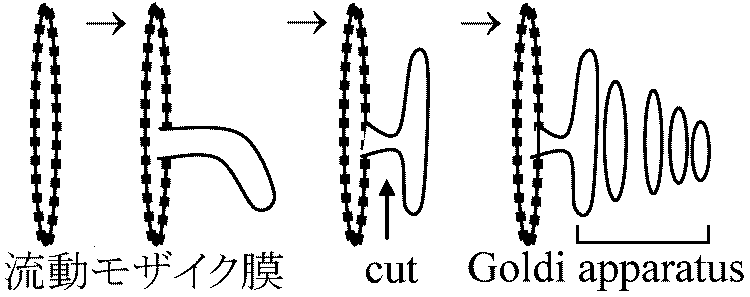
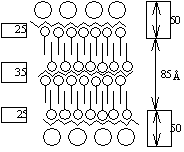
流動モザイク説 (fluid mosaic theory)1972 Singer SJ & Nicolson GL凍結劈開法 free-fracture: 膜を凍らせそれを裂き膜を調べる - freeze-etching 極性 polarity を調べるのに赤血球を使う利点: 細胞壁、核がなく膜だけの構造をとり観察し易い。これを低張液(0.14 M, NaCl)につけ溶血ghostさせる(等張ではvesicleができ、低張ではinside-outになってしまう)  膜成分である脂質は水和性のある極性端を膜両面に向け疎水性鎖を膜内面に向けた2重層を形成 → 2重層の厚さ7-8 nm
膜成分である脂質は水和性のある極性端を膜両面に向け疎水性鎖を膜内面に向けた2重層を形成 → 2重層の厚さ7-8 nm膜構成 protein: 多くは脂質2重層中にモザイク状にはめ込まれている。protein中の疎水性部分は膜内部に親水性部は膜表面に露出 → proteinには、この表面に露出した部分に結合しているものもあるし脂質の2重層の中に埋まるのもある 脂質2重層: 常温では結晶と液体の中間状態(= 流動性) → 膜内部に存在するproteinはこの流動性を持った脂質層の中をかなり自由に動くことができる。このモデルは細胞膜が動的な構造であることを示すと共に安定した構造であることも説明できる。この構造は細胞膜だけでなく細胞内種々の膜構造でも基本的な構造である 細胞膜成分にはlipid, proteinの他に多糖類がある。多糖類の種類は細胞の種類によって異なり細胞はそれぞれの表面に特有な多糖類を持ち、細胞表面多糖類の重要性が判明している 機能 function1) 細胞内環境維持伸縮性: 修復・再生・隔合維持: バクテリア・ビールス等の侵入を防ぐ(植物の場合更にcell wallがこの機能を果たす) 糖衣glycocalyxが働きの主 Takebe(建部)
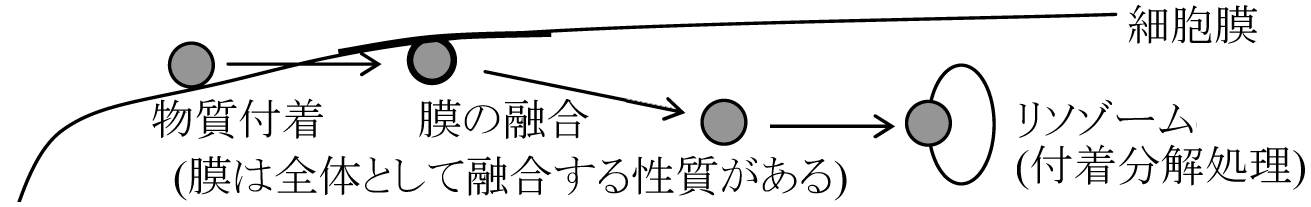
TMVより原形質体protoplast抽出 2) 物質透過・輸送a) 受動輸送 passive transport拡散による透過: 溶媒としての水はかなり自由に通す neutralな物質(no charge)であるのが膜の目の大きさpore sizeを考える時の前提 → chargeがあると小さなものでも通し難いものがある |
形質膜は巨大な脂質なども取り込む - 自由に透過 原形質分離 plasmolysis 外部濃度調節に蔗糖、尿素を用いて比較
物質: 分子量 濃度
蔗糖:_342 内 < 外 蔗糖では永久に原形質分離plasmolysis状態であるが、尿素では時が経つと内外部とも濃度が等しくなる(原形質分離が見られなくなる) → 形質膜にはある大きさ(尿素)を通す程度の孔がある b) 能動輸送 active transport拡散による物質移動のほかに、呼吸によって生じたATPを用い積極的な吸収や排出を行う現象 ナトリウムポンプ
エネルギーが必要: ATP →ATPase (energy) → ADP + Pi
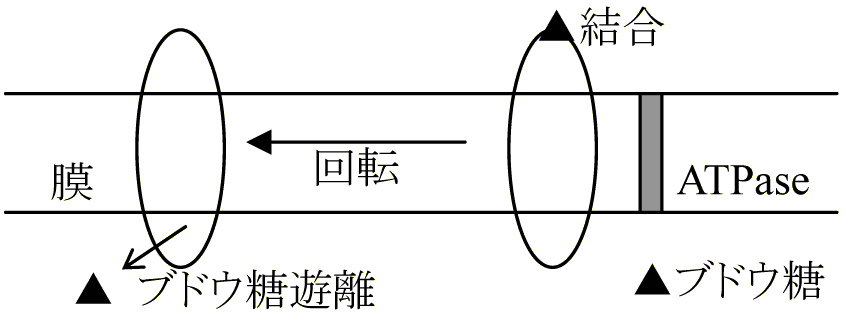
→ エネルギーを得るにはATPとATPaseが必要 選択的透過性 selective permeability細胞に必要な物質を選んで通す性質物質取込、排出にエネルギー(ATP)必要
認識タンパク質: ブドウ糖結合タンパク質、アラニン結合タンパク質、硫酸結合タンパク質
赤血球 27.4 K+: 細胞内 > 外: 血漿 5 mM 3) 情報伝達Con A (concanaralin A): 細胞を凝集させる働きがある→ 白血球に与えると白血球分裂が盛んになる → 細胞分裂促進剤 → DNA複製は核中 膜にCon Aと結合するreceptorがありCon Aを加えると結合し核にDNA複製情報を連絡している |
= 原形質膜 protoplasmic membrane, 古語
|
ほぼ半透性で選択的透過性
H2O, O2, CO2 化学組成: 脂質 lipid, タンパク質 protein 脂質: 脂肪酸グリセリド(fatty acid glycerides, 脂肪酸とグリセリンの化合物)。C17, C19類が多い
食胞 food vacuole (食細胞 phagocyte)食作用 phagocytosis: 固形分取込 → 器官 = ファゴゾーム phagosome他種細胞を取り込む ☛ 細胞内共生 飲作用 pinocytosis: 液体成分(コロイド)取込 → 器官 = ピノソームpinosome
取込過程 細胞基質 cytoplasmic matrix無構造 → 原形質流動タンパク質・酵素・RNA含む → 収縮性タンパク(アクチン・ミオシン) 細胞の相互認識培養層に種々細胞(肝臓細胞、腎臓細胞)をばらばらにし混合→ 培養層内でお互いを認識し同種細胞が集まり結合 |
認識能力は細胞膜表面にある 糖タンパク: 一種のアンテナ = 認識能力部分
→ この部分をぺプトン等で分離すると異種タンパク質も結合 生体内脂質脂質は生体膜主要成分
______________血漿_____牛乳 細胞表面の被覆・防護 → 高等植物葉、昆虫殻、皮膚、細菌細胞膜等 細胞中の脂質成分 貯蔵・運搬
エネルギーの貯蔵物質として脂質は高エネルギー分子なので適している 赤血球膜 神経細胞膜 → すべて細胞膜構造をとる 神経細胞末端 (筋肉細胞を含む)
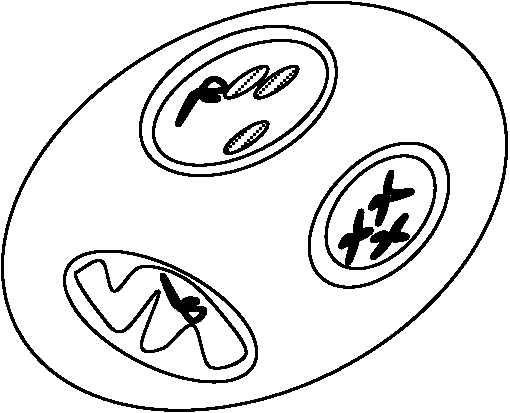
神経細胞末端と筋肉細胞はneuro-mascular junctionという形で結ばれる 核膜(eukaryotes, prokaryotes共), 小胞体, ミクロゾーム, ミトコンドリア, 葉緑体 |
|
= 細胞内膜系 細胞膜起源。網目状に細胞膜が発達したものであるが機能は細胞膜と異なる 膜を持った構造体。ER形態から細胞の種類、代謝活性の度合が判断可能なほど、膜系形態、量は様々。内膜系の役割は、細胞交流の場を提供し、合成産物を分離し、生産物を細胞内外に移送する通路となるものといえる
Ex. 膜タンパク質: イオン吸収 - Na, K, Ca = カチオン輸送、-SO, -PO4 = アニオン輸送 粗面小胞体(rough ER, R-ER), 滑面小胞体(smooth ER, S-ER)は機能分化が表れたもの 粗面小胞体 rough-surfaced endoplasmic reticulum, rER分泌タンパク質 → ribosomeがproteinを作り細胞膜にすぐproteinを渡し流動モザイク構造に使用 |
滑面小胞体表面にribosomeが付着しprotein(主として分泌タンパク質)の合成の場であるribosomeの状態によって遊離型ER、結合型ERの2種類がある 諸種物質合成の酵素を有する Ex. 脂質合成酵素により脂質を合成し、流動モザイク構造に脂質を渡す ribosomeは細胞膜表面に結合するproteinが存在すると小胞体に結合するミクロボディ microbody一枚膜: 直径0.2-1.0 μmの袋滑面小胞体が特殊化したもの。パーオキシダーゼ peroxidase (パーオキシゾーム, peroxisome)と呼ばれる酵素とグリオキシゾームglyoxisomeと呼ばれるグリコオキシダーゼを含む袋 |
 チュブリンが管状となり液胞近くに多量存在。微小管配列変化が細胞自体の移動・細胞内物質移動に関係
チュブリンが管状となり液胞近くに多量存在。微小管配列変化が細胞自体の移動・細胞内物質移動に関係チュブリン tublin: 微小管(細胞骨格)構成基本単位となるタンパク質 中心体 centrosome (中心粒 centriole)= 微小管形成中心 microtubule organizing center, MTOC動物細胞 (植物にはない) 微小管で構成される |
鞭毛 flagellum藍藻、紅藻、菌の一部以外は胞子に鞭毛を持つ鞭毛の種類鞭毛糸: 9 + 2 = 11 |
= ゴルジ装置 Goldi apparatus、ディクチオソーム dictyosome

構造 structure動物・植物に広くみられる単位膜の系膜表面にribosome付着せずS-ERに近い。内膜と連続している所が観察されることもあるが不連続である場合が多い。平板状の胞が平行に重なったもので、胞の縁には直径400-800Åの小さな胞球がたくさん付随する。胞球は平板状の胞がちぎれてできたもので溶液状あるいは粒子状の分泌物を含む ゴルジ体構造は3要素からなる
ゴルジ膜: 偏平な嚢 機能 function細胞分画法で単離したゴルジ体 → ガラクトシルトランスフェラーゼ活性顕著 → 糖代謝と関係深い |
植物細胞: サラニペクチン, hemicellulose, cellulose等の多糖を合成し、これらを細胞外に分泌し細胞壁新生・伸長・肥大に寄与。炭水化物を合成し分泌する細胞では少なくとも一部はゴルジ体の酵素により合成され、そういう炭水化物はゴルジ体部分においてその細胞の他の部分で合成されたタンパク質と複合体を作る リボゾーム上で合成されたタンパク質は粗面の内膜系を通じてゴルジ体に送り込まれ濃縮あるいは荷作りされて細胞から外に出易い形になる 脂質代謝においても一役果たす。精細胞が成熟して精子になるときの変態においてもある役割を成す リソソーム(リソゾーム) lysosome原形質成分を分解する酵素を含むゴルジ体に似た球状の小胞細胞が摂取した粒子や不必要な物質をゴルジ体からの消化酵素によって分解消化する 植物: 液胞がこの機能をもつ(リソソームがないわけではない) [植物]1次リソソーム: 加水分解酵素有 → 2次リソソーム: 1次リソソーム + ファゴソーム → 不要物分解(酵素による) |
|
= 粒子体、コンドリオソーム、スフェロプラスト 真核生物にみられ、幅約0.5 μmの糸状・顆粒状細胞小器官 主要機能呼吸 → TCA回路及び電子伝達系の主要酵素持つ= 好気的条件下の細胞エネルギー生産の場 mt固有のDNA, RNA存在
自信が複製・転写・翻訳の独立した機能を持つ
ATP, ADP: アトラクチロシド (atractyloside)により阻害受ける 構造高濃度のリン脂質とタンパク質を含む
→ 周囲の細胞質より屈折率高い → 生細胞でも位相差顕微鏡で観察可 → ATP合成の盛んな細胞においてmtが数においても構造においても発達していることが予想される 構造様々: 団子状から線状のものまで 大きさ様々: 幅0.4-3.0 μm, 長さ0.3-0.8 μm クリステ構造 crystae = -二重膜構造 二枚膜構造 = 外膜 outer membrane (細胞膜あるいはERに近い) + 内膜internal membrane (クリステをなす膜)
膜間層 礎質内に存在するもの
mt-DNA: 核DNAと分子の性質が異なる 二重膜外側膜は核DNAにより作られる。内側膜はmt内部のDNAと一部は核DNAにより作られる(重要なものは核からもらうがmtを作る遺伝情報は核にはない)1960 Sjöstrand
膜が2枚集まった2重膜であることを発見
calf heart mascleを低張中で重金属によるnegative stainingする mt全重量に対する割合 = タンパク質・脂質・他 (50) + 水 (50) mt全乾燥重量に対する割合 = タンパク質 (70) + 脂質 (25-30) + 他 (≤ 数%) 全タンパク質に対する割合 = 可溶性タンパク質 (20) + 不溶性タンパク質 (80) 脂質に対する割合 = リン脂質(不溶性タンパク質と結合) (≥ 90%) 不溶性タンパク質 = 構造タンパク質 + 電子伝達系タンパク質 表. ミトコンドリアクリステ長総和とチロクローム濃度の比較ミトコンドリア 外膜長 クリステ長 クリステ/ クリステ Cyt. a濃度b (μm)a 総和(μm)a 外膜 ダイコクネズミ 心臓 2.83 7.37 2.49 34 0.66 肝臓 2.62 1.99 0.70 12 0.18 ウシ 心臓/肝臓 1.08 3.70 3.56 3.7 肝臓 3.32 0.86 0.26 6 0.08
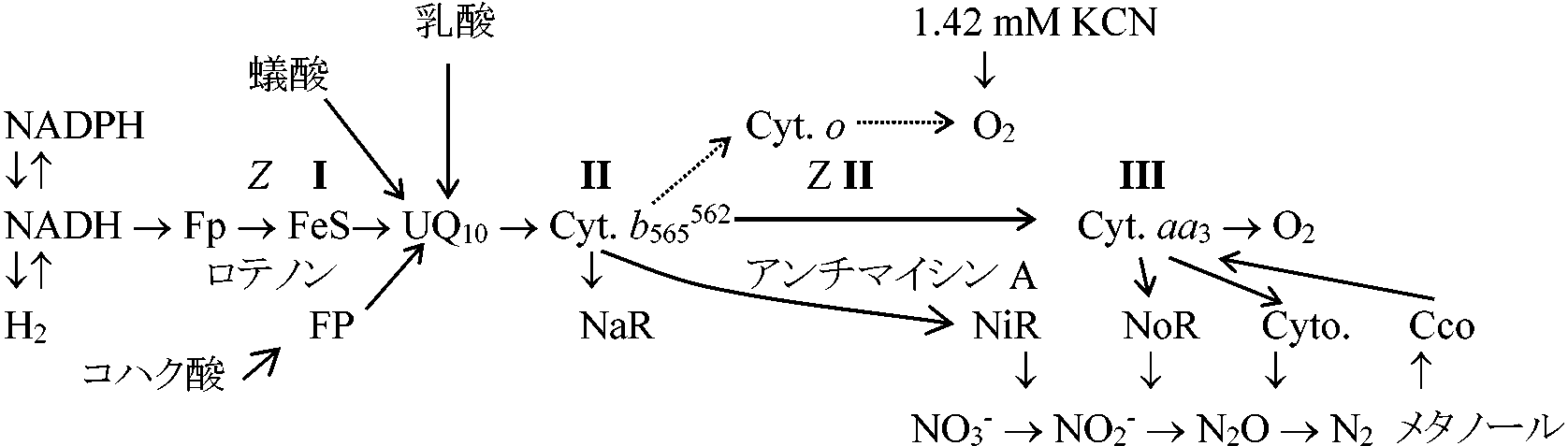
a. 1ミトコンドリアあたり平均長、または数(過マンガン酸カリ標本) 機能エネルギー生成系酸素呼吸 = ATP合成 (ATP synthesis)
C6H12O6 + O2 → 12H2O + 6CO2 + 38ATP (688 kcal) |
TCA回路 = マトリックスに存在 ⇔ 電子伝達系 = クリステに存在 CH3COOH → アセチルCoA 基本粒子(elementary body) = ATP生産部位 → ATPosome(ATPオゾーム) = F1粒子 ミトコンドリア結合時はATP合成、分離時はATP分解 = ATPase (ATP分解酵素) → 共役因子: protein ATP → [ATPase] → ADP + Pi 酸化的リン酸化
ミトコンドリアは水吸収・排出(ATPも一緒に出入)により収縮膨張を繰返す
現在否定
クエン酸 (citric acid) → トリカルボン酸 (tricarbonic acids)の重要性 - 酸素消費のために重要
→ 無機リン酸は増加物質の複合体となると考えた シアン化合物を加えると活動鈍る → シアン化合物はO2阻害を行うためO2の必要性が示唆された → ATP生成にO2必要: ADP + Pi → ATP 1940 Green, David Ehomoganated cells: 呼吸の活発な遠心分離部に酸化的リン酸化(oxydative phosphorylation)の中心組織を見出しcyclophoraseと名付けた → cyclophorase system 1945-50 Lehninger: mitochondriaを単離(isolation)する
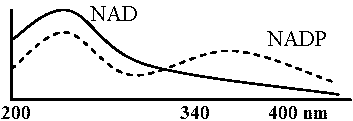
NADの還元により紫外線吸収波長変わる
TCA回路炭水化物・アミノ酸 → CH3-C(O)-COOH (pyruvic acid) → acetyl Co A__________________________________________CoA
オキザロ酢酸 → クエン酸 小胞 vesicles= 亜ミトコンドリア粒子無傷の小胞構造がエネルギー保存過程に必要 抽出すると外側の膜が外側になってright-side-in得られるが、一部は膜が逆転しright-side-out得られることがある
人工的にmitochondriaをばらす-亜mitochondria(vesicle)粒子化
トリプシン trypsine, 尿素 ureaで分解ののちに遠心分離
沈殿: 電子輸送行うががADPリン酸化できない分画(電子伝達系) |
色素体 plastid葉緑体は色素体の一種 葉緑体 chloroplast → 葉緑素chlorophyll
褐色体(褐藻・珪藻) phaeoplast → 褐藻素 fucoxanthin 白色体 leucoplast: chlorophyllを含まない アミロプラスト amyloplast: 葉緑体で生産された澱粉を貯蔵する プロプラスチド proplastid: 未発達色素 光合成色素表. 光合成色素の特性 (Govindjee & Braun 1974)色素: 吸収極大, nm (測定部) クロロフィル類
a (青緑): 435, 670-680 (生体内)
α-カロチン: 420, 440, 470 (ヘキサン中)
ルテイン: 425, 445, 475 (エタノール中)
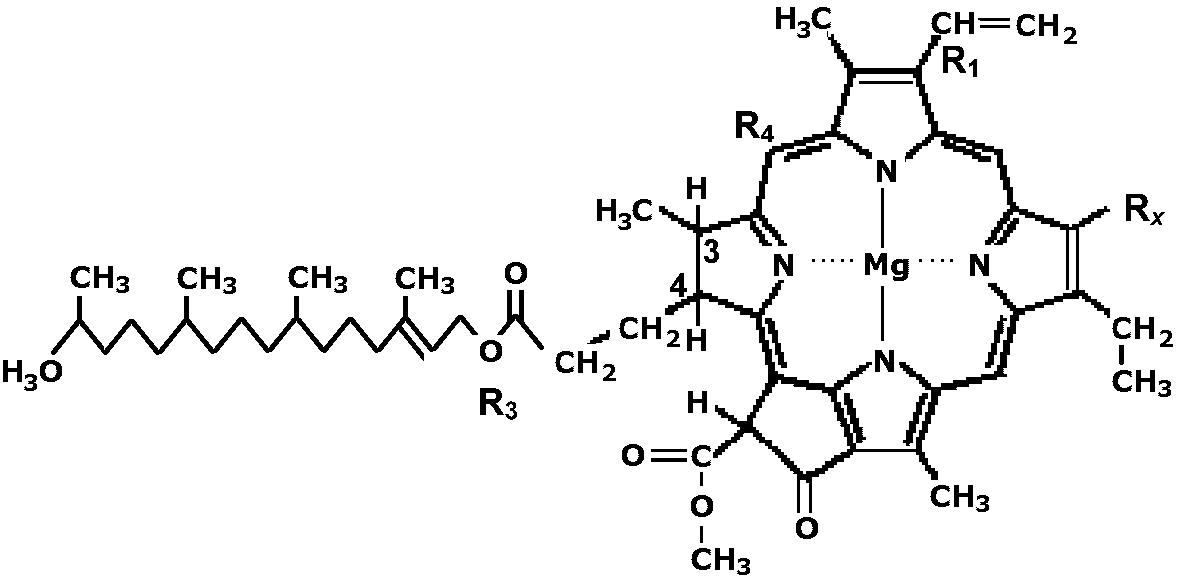
フィコエリトリン: 490, 546, 576 (蒸留水・生体内) [ 葉緑素抽出 ] 葉緑素(クロロフィル) chlorophyll, chlクロロフィルa:b比 (chl a:b)一般にchl a:b = 3:1 a:bは、同一種内のPmax/葉緑素比と相関 → 光合成効率(光化学系のアンテナサイズ)を決める要素 Ex. 同一種内では陽葉ほど大 Ex. chl a:b低下: 光化学系II(PS II)にある2つの反応中心のうち、光化学系II集光性クロロフィルタンパク質複合体(LHC II)を多く結合するαセンターの相対的増加 → 被陰環境への順化応答 クロロフィルとヘモグロビンは構造が似る → 共にポルフィリン化合物
ピロール核 中央元素 フィトール基
クロロフィル 4 Mg 有
ヘモグロビン 4 Fe 無
クロロフィルa, bと各種バクテリオクロロフィル(a, b, c, d型)の構造的関係 (= 分類基準)
|
表. R1, 3, 4, R2, R3, R4の結合基 R1 3, 4 R2 R3 R4 Chlorophyll a -CH=CH3 - -CO-CH3 phytyl ester (C20H38O-) -H b - * - - - Bacteriochlorophyll a -CHO-CH3 Dihydro -CO-OCH3 phytyl ester -H b - - 構造未知 - - c -CH-CH3 - -H farnesyl ester (C15H25O-) -CH3 d -CHO-CH3 - -H farnesyl ester - *: 3: CH3がCHOに置換。4は上と同 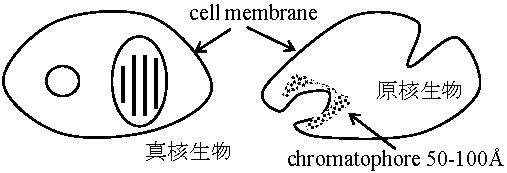
構造原核生物 (prokaryote)ある程度は構造分化があるが、色素胞 chromatophore (藍藻の場合)または特別な組織なし→ いってみれば葉緑体 = バクテリア個体全体 幾つかの構造を取る ex. nucleoid カロチノイド等が結合(associate)している Chlorophyll a chlorobacteria 緑色硫黄細菌 rhodobacteria 紅色硫黄細菌 H2S - bacterio chlorophyll 真核生物 (eukaryote)クロロプラスト chloroplastchlorophyll a, b, (c, d) 平均的なものは5 μm、厚さ3 μmのもの。葉肉の細胞には数10個含まれる 2枚膜 = 外膜(envelope, or external membrane envelope) + 内膜(internal membrane) マトリックス(基質, 礎質, matrix): 溶液状態 内膜 internal membrane チラコイド thylakoid: 内膜がmatrix中へ侵入した薄い膜の袋 – 膜数が分類基準となる グラナチラコイドgranum thylakoid
グラナ granum (pl. -a): 葉緑体内でチラコイドが層状に並んだ部分 ラメラ lamella: 葉緑体膜に包まれた内部、層状構造 グラナラメラ grana lamella: グラナgrana部分にあるlamella → 光合成(明反応)を受ける色素局在 ストロマラメラ stroma lamella: ストロマ部分にあるlamellaEM観察葉緑体外側に厚さ15-20 nmの葉緑体膜 = 3層構造: 高電子密度の2枚の膜間に低電子密度の膜ラメラは葉緑体を切断した断面につけられた名だが、切片で層状構造にみえるのは立体的には偏平な薄い袋(膜胞状のもの)である。この袋とチラコイドthylakoidの間には大小2種類があり、大thylakoidの間に小thylakoidが積み重なり密着している。小thylakoidは円盤上で数枚-数10枚が密に積み重なる。これがgranaでgranal lamellaはその様な重なった円盤の断面をみたもの。大thylakoidは基質stroma部分に伸びる。この断面がstroma lamellaである。 緑藻等の藻類葉緑体はgrana lamellaとstroma lamellaの区別がなく、thylakoidは1種類。緑藻や接合藻類等の葉緑体中には1-多数の光を強く屈折する小体がある。この小体はピレノイドと呼ばれ周囲にデンプン粒がみられデンプン形成に関与しているとみられる 基質中には同化デンプン粒、ribosome等の顆粒、DNAの微繊維等もみられる |
|
RNAとproteinからなるタンパク質合成の場
組成: RNA 60%, protein 40%の種多 リボゾーム粒子 = だるま状構造
____小亜粒子 small subunit_ 細菌型と真核細胞型細菌型: 前核細胞生物 (細菌, 藍藻, mitochondria, 葉緑体) 起源S = 70 直径 = 140-210Å 2.7 × 106 (RNA = 65%, protein = 35%) 真核細胞型: 核 → 細胞質細菌型ribosome 真核細胞型ribosome S 70 80 直径 140-210 Å 220-300 Aring; 2.7 × 106 4 × 106 (RNA = 65%, proteins = 35%) (RNA = 45%, protein = 55%) 小亜粒子 大亜粒子 小亜粒子 大亜粒子 M.W. 全体 0.9 × 106 1.8 × 106 1.3 × 106 2.6 × 106 RNA 0.6 × 106 1.2 × 106 0.6 × 106 1.7 × 106 S値 全体 16S 23S, 5S 18S 28S, 27S, 5S RNA 30S 50S 40S 60S タンパク質 糸状 糸状 数 21種* ≥ 34* ≥ 24 ≥ 34
* 順序良く結合 -----------→ 原核生物 ------------→--→ 真核生物 --→ 70S =================⇒ 70S =(小器官内)=⇒70S (ミトコンドリア・ ⇓ 葉緑体リボゾーム) リボゾーム 80S変異 ⇓ 80S = (細胞質内) =⇒ (細胞質リボゾーム) |原核細胞の起源| |真核細胞の起源| 図. リボゾームの原核型(70S)から真核型(80S)への進化 |
再構成 (E. coli)リボゾーム ribosome を2つの亜粒子に分離するにはMg2+濃度を低くする。濃くすると再び結合する小亜粒子 30S: S1-S21 → 13種pK > 11、19種 > 7, 30S構成に8種必須(常在多) 再構成: 16S RNAとS1-S21を精製し再構成順序を明らかにする Ex. 16S → S4/S8 → S7 → S19/S9 → S14 → S10 → S3 機能: S12: 誤訳(曖昧)因子。不在時に正しい翻訳, S11: 正訳因子。S6, S15, S18も関与, S7: 調節因子, S13: m-RNA結合 大亜粒子L1-L34は4種を除きpK > 7, 50S粒子1個あたり複数分子存在するものもあり
再構成: 23S RNA + L1-L34 + 5S RNA → 50S ポリゾーム polysome or polyribosomem-RNAにribosomeが複数結合したribosomeの総称 ⇔ 1 ribosome = モノゾーム (monosome, or monoribosome)polysomeはprotein合成時型で、mRNAは小亜粒子に結合。mRNA始めの部分にribosomeとの結合部位 Ex. ヘモグロビン作る際のpolysome = mw 64500 = アミノ酸(α鎖のみ) 141個
1アミノ酸生成に3塩基配列必要 → 141 × 3 = 423個の塩基からなるm-RNAが最低でも必要 |
|
原則: 細胞は1個の核を持つ → Def. 細胞核 = この1つの核
無核細胞もある Ex. 赤血球 外側は核膜で囲まれる。核膜は内外2枚の膜(単位膜)よりなる。核膜の所々に小孔あり、小孔を通り核と細胞質の間に物質交換が起る。核膜外側にはribosomeが付着することが多く、核膜はしばしば細胞膜に突出し小胞体の膜と接する。核膜は成分や構造から小胞体膜と相同のものと考えられる 核膜 nuclear membrane2枚膜 ER 外膜内膜 膜間層 perinuclear space 膜孔 nuclear pore 染色糸 chromatin: 核小体nucleolusがついている 核質 nuclear substances: 染色糸 + 核小体 核液 nuclear sap 仁 (核小体) nucleolus, pl. nucleoli核分裂前中期-後期を除くほとんど全ての細胞核内に存在する小球体仁数は通常生物種によって一定だが、同じ生物でも組織によって形・大きさ・数が異なる。この変異性は細胞活性時の蛋白質合成能と関連している
Allium cepa (タマネギ): 1仁/1細胞 protein → 重複: 一遺伝子から多数のm-RNAをつくりproteinを合成 仁中RNAはpre-RNA (precusor r-RNA) → 仁はribosome合成の場 仁主成分: prorinで染色、Rnaseで消化 → RNAとproteinsの結合したもので基質はproteins(80%以上) 1957 Allfrey, Mirskey, & Osawa: 核自体がprotein合成すること確認 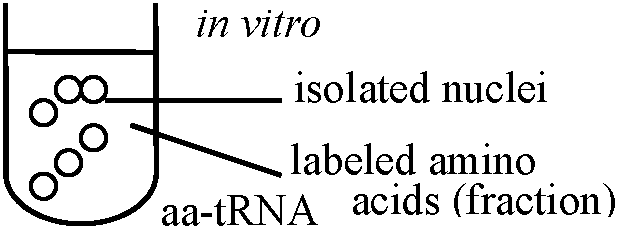
amino acidのままなら沈殿しない acid insoluble fraction → protein合成は行われた
Pronin (RNAを染める) → nucleolusがpink / Methylgreen (DNAを染める) → 核質がvioletと染め分けできる 成分1950 Caspersoncondensed chromatinをnucleolus associated chromainと命名 1952 Vincent
ヒトデ卵細胞を低張液に浸し加圧し核を取出し、更に遠心で核小体を取出す(但し低張液だと水に溶け出すので分画法発達すると等張にする) →
核DNAと核小体を囲むDNA(G, C rich)では組成が異なる
DNA/RNA hybridを作ると、このDNAは外のDNAとつかず核小体のDNAのみにつく
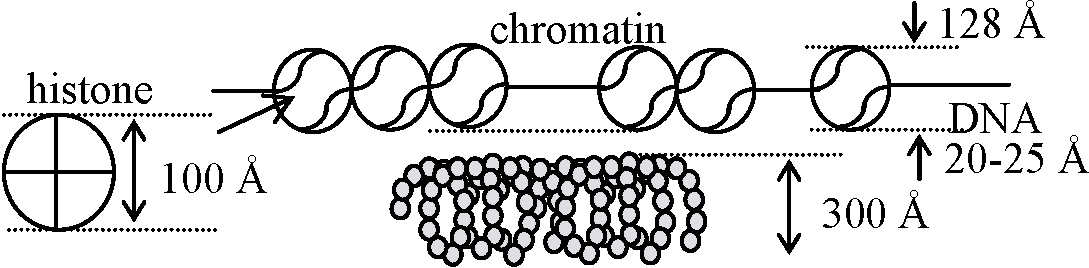
→ r-RNA geneの存在箇所を確認 核質 nuclear substanceschromatintin, nuclear sap, nucleolus, nuclear skeleton, nuclear vacuoleダイコクネズミ肝臓 0.25 Mショ糖に浮遊させ音波処理 = 核破砕 → 遠心, 2500 × g, 8 min. → 沈殿 [上澄 (染色質、核液)] → 0.88 M ショ糖に浮遊 → 遠心 → [上澄] → 上の処理をもう一度繰返す → 最終の沈殿 = 核小体 ヒストン histone塩基性色素で染色される構造体
クロマチン: 分散状態 - 休止核 H1, H2A, H2B + 2本鎖DNA分子が結合 20-25 Å, H3, H4 → 2H2A 2H2B 2H3 2H4: histone分子8個が結合しそれにDNA鎖を巻き付けたものがchromatinとなる クロマチン/染色糸/染色質 (chromatin)= DNA + protein + RNA。核小体nucleolusがつく1928 Heitz: ゼニゴケ細胞をカーミン染色 異常凝集部heteropycnosis発見: heteropycnosis数 = chromosome数 → pro-chromosomeと考える |
1940 Darlington & La Cour
Feulgen staining (for specific DNA) was made after Trillium is chilled at 0-4°C for 4 days
HCl or NaOH, NaCl, 60-65°C処理で染色体は殆どstainしない 二本鎖の螺旋構造: DNA: (r) +, (l) - 真核生物核内に存在する好塩基性物質。現在はDNA塩基性核蛋白質(複合体を主成分とし非ヒストン蛋白質)および小量のRNAを含む集合体に対して用いる。細胞サイクル各期、遺伝的活性化・不活性化状態で構造が著しく異なる。heterochromatinとeuchromatinに大別される 異質クロマチン condensed chromatin (heterochromatin): 分裂期間中も見え続ける凝縮部位 heteropycrosis このDNAがRNAに転写されないため遺伝的不活性染色体上でhetero-chromatinになる部位は決まっている。Heterochromainはminor chromatinでありアミノ酸情報には関与しない
真性クロマチン diffused chromatin (euchromatin): 分裂時に見えなくなる解けたクロマチン。遺伝的活性部分 ヌクレオゾーム nucleosome110Å = 200 base pair DNAがhistoneの適当な基と結合したもの
2分子のH2a, H2b, H3, H4を含む (1 × H1も含む) 直径 = DNA直径 × 7, 太さ = 1/7
ソレノイド solenoid : chromatin (nucleosome)が螺旋状となる ↔ ヌクレオソーム (100 Å) ↔ DNA 2重螺旋 核液 nuclear sap or karyolymoph無機塩類、低分子物質等が考えられる – 未詳
soluble protein (free enzyme), nucleoside, ヌクレオチド分解酵素nucleotide
核からRNA + proteins, rDNA (r-RNA gene DNA) + proteinsを取り除くとprotein netが残る 核の糸 (thead)1953 Brinstiel & Hyde: Peaのchromatinを電子顕微鏡観察 → 35-45Åの糸1965 Du Paw
海面展開法: ミツバチの胚 (chromatin, chromosome)
1974 Clark & Felsenfeld, Axel et al.
Chromain + DNase II (staphylococccal nuclease) → DNA extract → analysis/results |
|
(Brachet & Lang 1965) 再生 (regeneration)カサノリ Acetabularia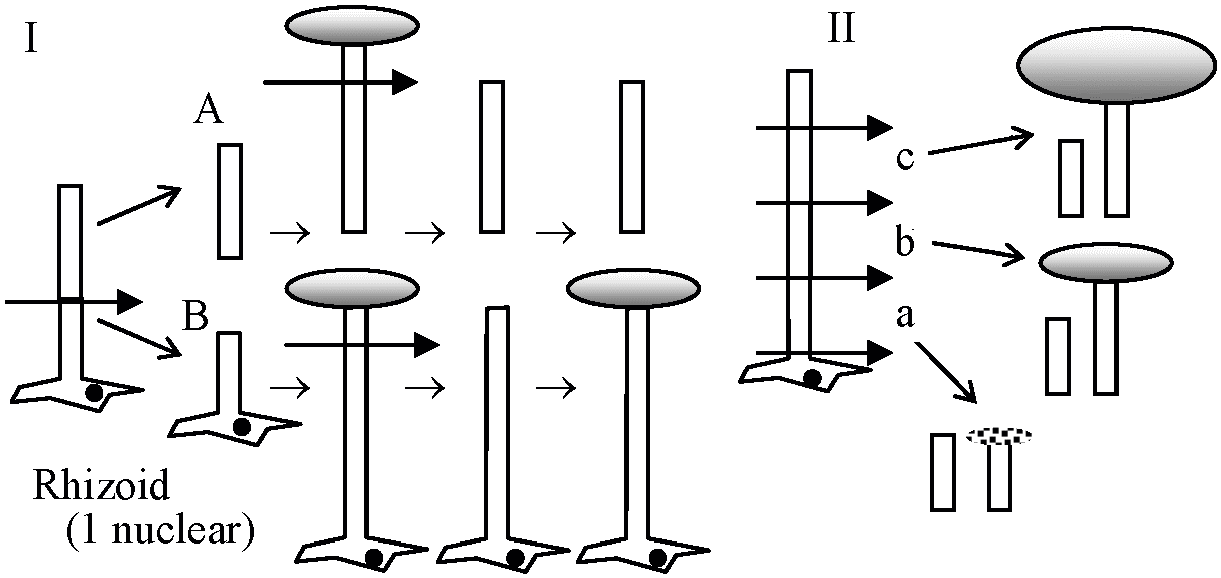 I: 無核片(A)は成長し傘再生するが、できあがった傘を切り落とすと2度目の傘再生は起こらない。有核片(B)の場合は何度でも傘の再生を行う → 再生には核が必要 II: 茎の部位による再生能力の差。頂端に近いほど再生能力が高い → 傘の部分に傘形成cap formationに関する物質がある – 核に支配されているが依存の程度は低い 傘capが発達すると核が急速分裂(1万-1.5万) → 急速に傘へ移動 → 嚢果cyst → 減数分裂 → Zoospore 移植A. mediteranea (med)とA. crenulata (cren)の移植 – 傘の形態で区別可能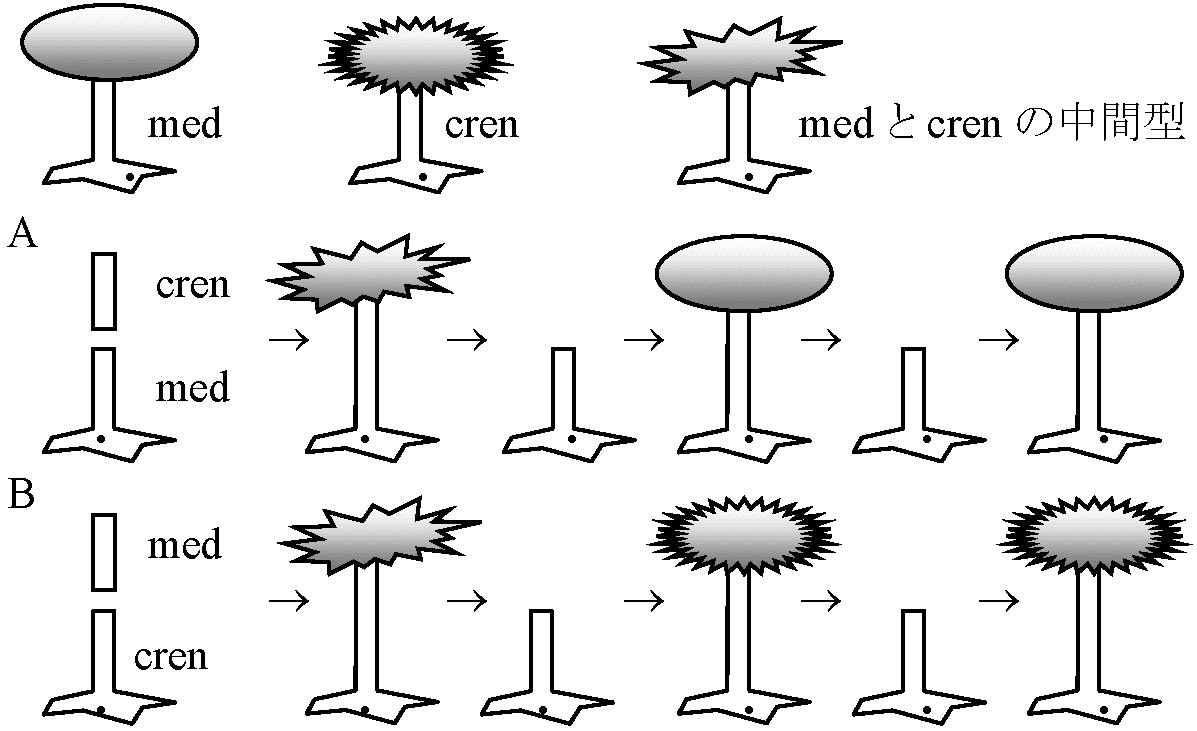 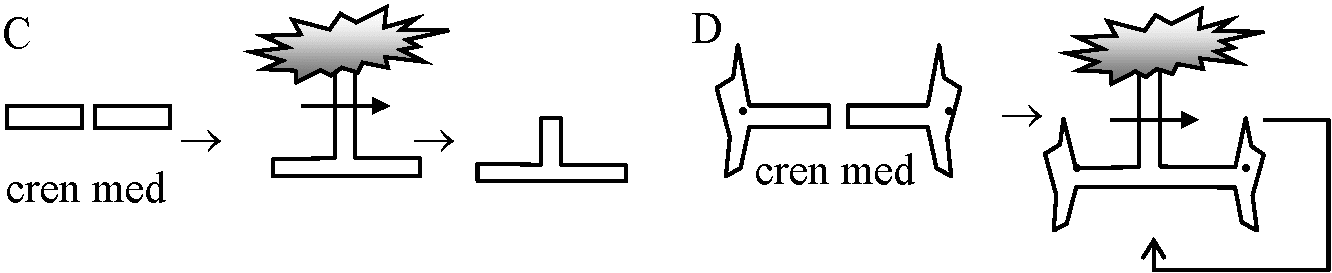 |
1959 Werg G
1) Carnoy's fluid (for fixation) + Azorcarmine (for staining) 
growth point → stainingに差
2) Azur I (for staining RNA) → 1)と同じ結果を示す → RNase処理にsensitive → RNAもある
即ち、リボゾームにm-RNAが付いた状態で染色している 1958 Ohsawa & Hotta: 1次元成長から2次元成長への転換機構を調べる
G C W
1次元 13.5 7.2 12.3
2次元 12.9 10.5 9.2
8 azagnanine (RNA inhibitor) 13.5 8.2 12.0
Base ratioに変化が起こる: RNA inhibitorを入れると変化が起こっていない(現在データは否定的だが、考察は正しいと考えられる不思議な結果) 1965 Nakazawa & Tanno2, 4 cells → actynomycin処理(RNA合成阻害剤) → 2次元成長止まるが1次元成長進む → RNAが形態形成に関与していることは間違いない1963, 1964 中沢: Fucus - MP stain
Fucusで3H-uridineを取りこませ追跡すると同様の結果を得、中沢の実験追証 集まらせる要因・RNAの正体・分化の"前触れ"現象 Cf. 化学分化: peroxidase, indophenol oxidaseの細胞内での分化の違い = 重力仮説 |
[ 染色体遺伝学 ]
|
|
| 染色体観察 | |
|---|---|
ショウジョウバエ唾腺染色体観察Observation of salivary gland chromosomes in Drosophila目的Drosophila数種幼虫の唾腺染色体観察 - 染色体構造、変異等の理解深める材料 (特徴)Drosophila virilis 2n = 12, D. melanogaster 2n = 8, D. immigrans 2n = 8方法
|
観察ポイント
既成の永久プレパラート ヒト染色体観察
材料ヒト白血球培養細胞から得られた染色体永久プレパラート(白血球培養 → 増殖期にコルヒチン又はコルセミド処理 → 細胞を集めカルノア液固定 → エア・ドライ標本 → ギムザ染色 → 永久プレパラート)結果
|
| 染色体解析 | |
フローサイトメーターflow cytometer高い精度での核型分析flow karyotyping可能非常に細い流液中に細胞等の微粒子を高速度で流し、これにレーザー光照射し、微粒子に当たって散乱する光や、予め蛍光物質標識した微粒子が発生する蛍光を測定する装置の総称 |
フローサイトメトリー flow cytometry: その方法あるいは技術
Flow karyotypingによる染色体異常解析や植物染色体解析等も試み |
|
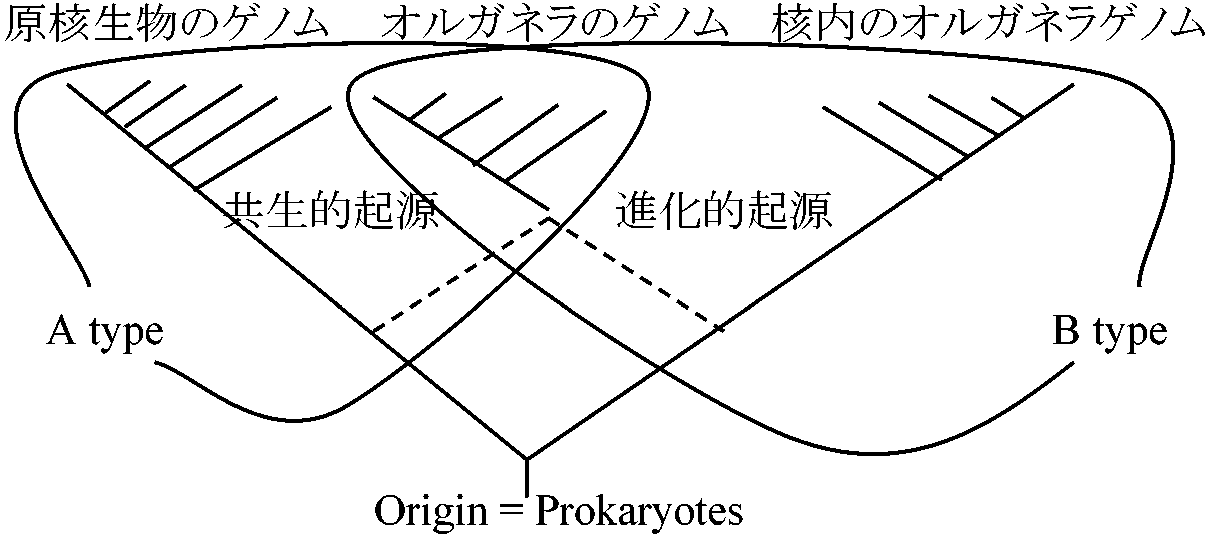
1959, 61, 62 Robertson: 3相説 = 古典的細胞像/共生説 → EM発達し否定 1962 Ris & Plaut: chlamydomonous葉緑体からDNA発見
chloroplast DNA - turnover (代謝回転): 二本鎖環状double strand circle - 40 μm (5-6分子/chloroplast) 同様にミトコンドリアにもchloroplast DNAと似た構造をとるDNAがある + リボゾーム顆粒らしいもの確認
↓ 寄生説(mitochondriaもchloroplastもbacteriaだった) 生命誕生後、原始生物は次第に新環境適応能力を獲得した。能力を捨てることもあるが(Ex. 欠失突然変異)必ずしも不利ではなく、外界からある物質が常時供給される環境に生育すれば、その物質合成能力を放棄しても差し支えず、むしろ、そのエネルギーを別部分に回せ経済的で有利 (Lowff 1943) → 葉緑体、ミトコンドリア共にDNA存在が確認された 1964 Lehninger: "The Mitochondrion"終章「ミトコンドリアの進化的起源」の節設けメソゾームの起源と共生説述べる 1965 Sitte Peter: "Ban und Feinban der Pflanzenzelle"
プラスチド由来と細胞区画: ミトコンドリア等の系の通則 共生過程 先カンブリア紀: 藍藻、高等植物型の光合成始まる = O2増加 → 既存生物の危機: 無酸素状態で生き延びる方法
被核細胞生物成立以前の生物
好気性裸核微生物 = pro-mitochondria (self-genomeによる調節 = Emden-Myerhoff反応) → 大形細胞体成立 + 裸核性藻類 → DNA量増加 ↓ 原生動物 → 有糸分裂の必要性 共生 = 被核性植物: protoplastidの取り込み(藍藻等にみられる) 1968 Roodyn & Wilkie: "The Biogenesis of Mitochondria"細菌とミトコンドリアの相似
核 裸核細胞 ミトコンドリア 被核細胞
核膜 無 無 有
2.5 nmフィブリル 有 有 (染色体)
ヒストン 無 無 有
染色体 無 無 有
分裂装置 無 無 有
電子伝達系の所在 細胞膜 内膜 (ミトコンドリア)
タンパク質合成
クロラムフェニコール阻害 有 有 無
アクチノマイシン阻害 有 有 無
RNase阻害 有 無 無
酸化的リン酸化依存 無 無 有
可溶性分画の添加 不要 不要 必要
アミノ酸のリポタンバク 有 有
質の優先的取り込み
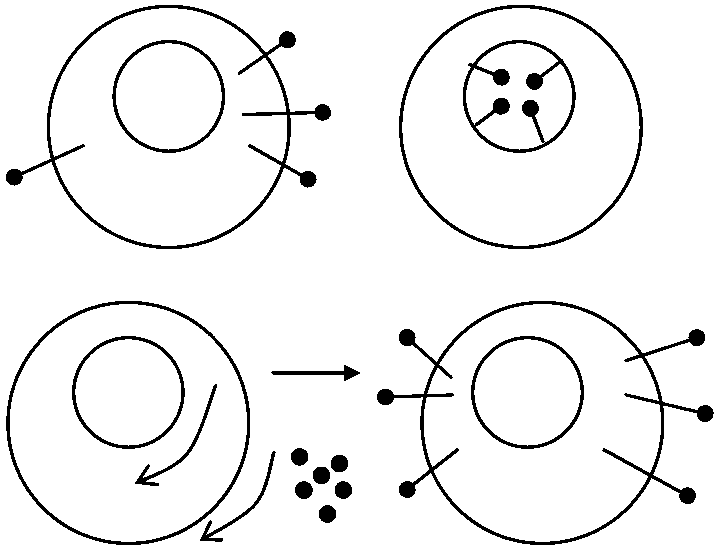
共生説(細胞共生進化説)かつての独立生物が退化し他細胞内共生 - 葉緑体やミトコンドリア形成
Ex. ゾウリムシ体内にクロレラの一種が共生 共生説論拠
マーグリス真核細胞共生説菌類起源の有力説 → 菌類は植物より動物に類縁の近い生物RNA塩基配列: 担子菌類 ≈ 褐藻 ↔ 子嚢菌類 ≈ 紅藻 藻類起源説: 従来の説。菌類は藻類に起源 ↔マーグリス説
担子菌類との共通祖先から葉緑体を細胞内に取り込み褐藻分化 ミトコンドリアとバクテリア
1962, 63 Nass & Nass: 融合コロニー説 colonial association theory 被核細胞生物の起源をSaganと比較し融合コロニー説提唱。弱点は宿主となった細胞の起源の説明を欠く 被核細胞生物成立の仮定 → 現在否定的
表. 藍藻と紅藻の類似性
1967 Raven 多元共生(重複共生)説
藍藻 紅藻 ミトコンドリアは多元か? – ミトコンドリアの不均一性 1969 Allsopp: 原始藻類説被核生物である藍藻から被核性の植物が進化した 紅藻が原核生物と真核生物の中間的性質を色々備えており、藍藻との類縁を偲ばせる唯一の真核生物 1969 Bell: 細胞膜陥入説
"Control of Organella Development" Symp Soc for Exp Biol London
現世藻類には葉緑素、葉緑体の構造で区別されるグループがある。それらは異なる光合成裸核生物を共生させた前被核生物の子孫である。また光合成裸核生物の大部分は死滅し藍藻だけが現存している 現世共生生物無関係に多発的に起こる1. 藻類が共生すると細胞壁退化現象が見られる Ex. Convoluta roscoffensis (動物)にPlatymonas convolute (珪藻)が共生 ⇒ 珪藻は細胞壁、鞭毛、眼点を失い、体を指のように長く伸ばすようになる ⇒ 宿主との接触面積が増す 2. 紅藻プラスチド1969 Alusopp: 藍藻細胞壁、プラスチド、紅藻葉緑体 = フィコビリン有する
分類上の諸形質は共通性乏しが、これら3群はフィコビリンを有する 4. 褐藻、ヒカリ藻、不等毛類、渦鞭毛藻類のプラスチド 1973 Lee 一元共生説: 進化を考える際の原則
I. 全器官、有機物、細胞内構造 –単系統的に考察したほうがよい?
グラム陰性菌 = 細胞壁薄い → 物質授受が用意(= 共生し易い)  図. Leeの系統樹
Raff & MahlerとUzzell & Spolskyの論争R & M: Sagan、Malguisらを批判
被核生物には好気性のものは極めて少ない。広い目で見れば被核生物は嫌気的なところで生存するものもあるが、好気条件での代謝産物を利用しているにすぎず、嫌気的被核生物は存在しないともいえる |
⇒ 被核生物 ≠ 単なる好気的オルガネラをもった嫌気的細胞質
表. 遺伝情報の細胞内分布: ミトコンドリアDNA, RNA、タンパク質合成系のタンパク質遺伝子所在 (mit: ミトコンドリア)
共生進化モデル (Raff & Mahler 1972)
タンパク質
機能特性
所在
この際、膜が閉じ酵素合成系の問題が生じる。そこでリボゾームおよび一部呼吸系のための遺伝子を持ったプラスミドをミトコンドリアに移した - 現実的ではない難題(目的論的) 1973.1.9 Uzzell & Spolsky: on Science
3.7 Raff & Mahlerへの批判 Saganの説は認めても良いが拘束力はない 被核細胞進化モデル (Uzzell & Spolsky 1974)
__↓ 被核細胞の元となる裸核細胞
⇒ ミトコンドリア、プラスチドなく、cytosisで珪藻・遺体等を食べ生存。膜状ER、ゴルジ体なし。リボゾーム少 1977 John & Whatley: P.p.自身は発酵のみ - 共生好気性細菌に依存し呼吸
⇒ ミトコンドリアの起源?は被核細胞成立の最初の段階を"Pelonoya stage"と呼ぶ
胞子はない。直径1 μm。土壌中好気性細菌
NaR: 硝酸還元酵素、NiR: 亜硝酸還元酵素、NoR: 亜硝化窒素還元酵素 共生系の進化 (Whatley et al. 1981)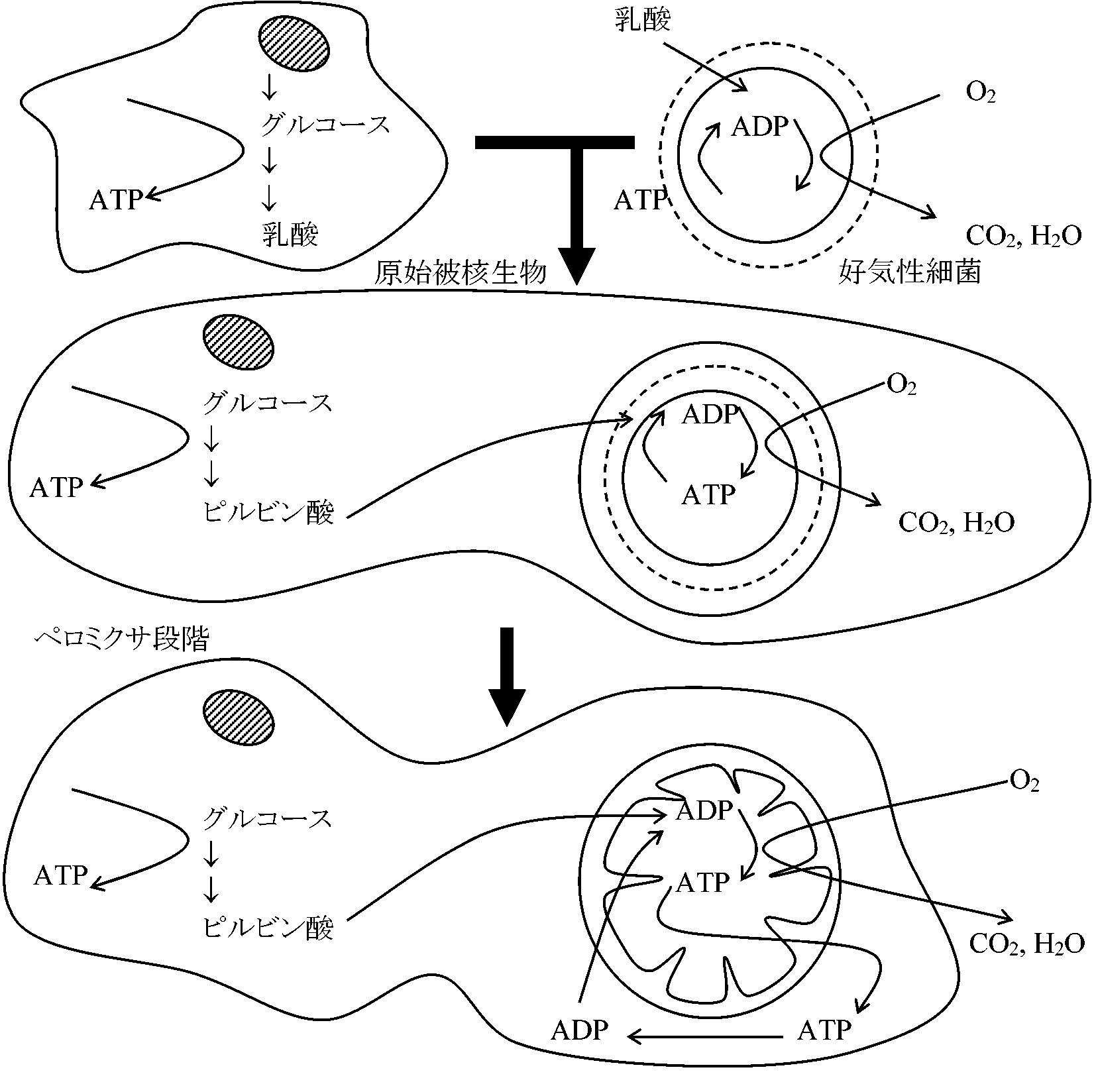
発酵性原始被核生物が呼吸をする細菌を取りこみ相助生活を始めた。そればペロミクサ段階発酵の最終産物が共生した細菌の呼吸基質となる。その後、アデノシンヌクレオチドキャリアーを得て細菌のATP合成能が宿主に役に立つようになりミトコンドリアを持った被核生物となった 緑色ホヤ green acidian緑色 = 藻類(藍藻)共生: 空家の宿主(ホヤ) aposymbiont 極稀⇒ 共生は脊索動物(ホヤ)という比較的高等なものに及ぶ Synechocystis didemni Lewin: ホヤ共生以外の生活様式ない
結論: このホヤ共生藻は典型的藍藻ではなく、Chl a, b有し緑藻に似るが、藍藻の特徴を有する (Lewinはこの生物を新しい門とした - 一般的に不採用) 昆虫細胞内共生 intracellular symbiosis in insects1887 Blochmann F: ブロックマン体 Blochmann body = 細菌1964 Lehninger: 共生体と細胞器官の類似を指摘 1965 Buchner P: 昆虫類130万種中の10%以上が定常的に共生細菌有する ホスト食性・栄養要求性 Ex. 肉食性のものは共生細菌少 菌細胞 mycetocyte: 共生細菌を収納する巨大細胞菌細胞塊 mycetome: 菌細胞が集合した特殊器官 共生微生物: 経卵伝播(共生球 symbiont ball) - 自由生活段階がないアブラムシ aphid28S rRNA, large subunit: 3500-4500 bphidden break (訳なし): 28S rRNAの途中でリン酸エステル結合中断(切目) gap deletion: 28S rRNA分裂時に一部の塩基が消失すること hidden breakを持つ種と持たない種がある
後口動物: なし 1978 Ishikawa (石川統): rRNAは共生微生物のものが主 ≠ hidden break 1982 Ishikawa: ゲル電気泳動: 複数のマイナーピーク- 共生体rRNA不安定 1984 Ishikawa: ナリジクス酸 nalidixic acid - 原核細胞DNA合成特異的阻害 単離共生体の3H-チミジン取込み阻害 = 共生体は原核細胞的 |
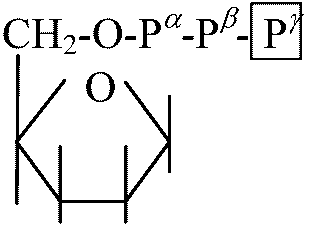
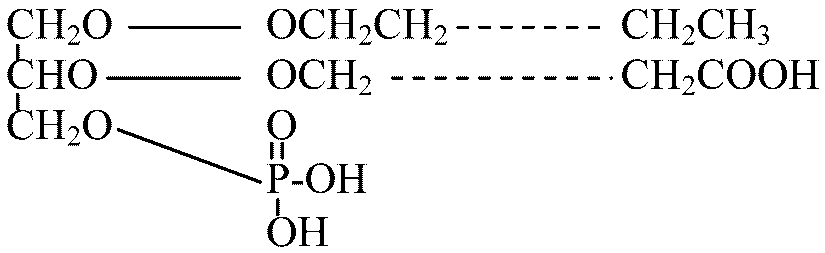


 →
→