(2025年12月2日更新) [ 日本語 | English ]
HOME > 講義・実習・演習一覧 / 研究概要 > 小辞典 > 植物解剖学
|
Def. 組織 tissue: 形態的にも機能的にも似た細胞の集合体
⇔ 複組織 compound tissue: 組織と組織系の中間的なもの ⇔ Def. 組織系 tissue system: 特定機能を果たす一連の組織の集合体
= 表皮系 + 維管束系 + 基本組織系 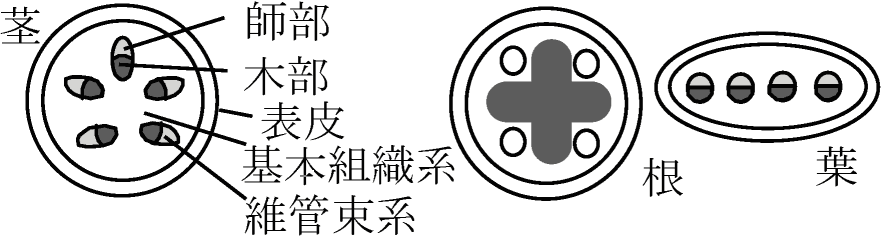 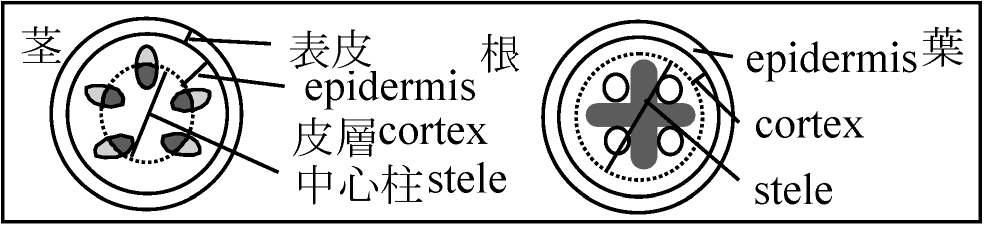 表皮系 (epi)dermal system Ex. 毛状体、根毛
表皮(細胞) = 外部覆い保護: 普通1層(2-複層もある) 双子葉植物 - 方向性弱い配列、単子葉 - 葉脈に沿い配列多
毛 hair: 多細胞・単細胞ある 基本組織系 fundamental system: 栄養貯蔵、光合成、支持構造提供
厚壁細胞(硬化細胞) stone cell or sclereid: 細胞壁が厚く硬い細胞
無限成長 indeterminate growth Ex. 茎頂、根端、形成層 |
分化位置による分類 [頂端分裂組織 apical meristem] 茎頂分裂組織 shoot apical meristem
その上に生じる多くの葉原基により隠されていること多 [側部分裂組織 lateral meristem] 維管束形成層 vascular cambium (= 形成層 cambium, s.s.) コルク形成層 cork cambium [介在分裂組織 intercalary meristem]
分化進行した部分に挟まれるが未分化のまま細胞分裂能を持つ部分 前分裂組織 protomeristem分裂組織の前段階 = 分裂組織中で未分化段階にある細胞集合体分化進むが細胞分裂能を持つ部分 = 前形成層 + 前表皮分裂組織 protoderm + 前基本分裂組織 前形成層 procambium: 木部師部等の一次維管束系細胞に分化
前形成層と基本分裂組織の分化は連続的に起こるため初期は区別困難 → 前形成層と維管束間柔組織を作る基本分裂組織が分化 茎頂で分化してから暫くは多くの細胞が細胞分裂能を有し成長する 分化過程求頂的分化 acropetal differentiation: 分化が基部から頂端部へ向かう 多くの前形成層は求頂的分化 (例外も少なくない - 特に単子葉植物) ⇔ 求基的分化 basipetal differentiation: |
|
Def. 維管束 vascular bundle = 木部 xylem + 師部 phloem
水や無機・有機養分の通導や、植物体の機械的支持に働く複合組織 Def. 維管束植物: 維管束を持つ植物 = シダ植物 + 種子植物通導組織 (通気組織, aerenchyma)[ 外側 ]表皮: edpidermis: 植物一次組織のうち最外部にある細胞層 (≥ 1)
原表皮 dermatogen, protoderm: 表皮を作る分裂組織
厚角組織 collenchyma: 厚角細胞から構成される組織 サフラニンに非染。細胞はある程度成長可能で、生細胞多
内皮: 維管束周囲の鞘や円筒作る基本組織層 中心柱師部: 複組織 = 師管/師細胞 + 師部柔組織 + 師部繊維
師部は木部に接する種多
繊維細胞 fibrous cell: 縦長・集合系 + 厚壁細胞: 細胞壁全体厚い 普通の細胞壁と違い、サフラニンに良く染まる(= リグニン)。堅い代わりに伸縮性なく死細胞多。形から細分類され、茎には縦に細長い繊維細胞 fiber cell (細胞壁厚く、全体が細長い形をした細胞)多い 師管(篩管): 篩細胞(管状構造とる) (+ 伴細胞) 分裂組織
木部 xylem: 複組織 = 導管 + 仮道管 + 木部柔組織 + 木部繊維
形成層細胞膜壁にリグニン蓄積 → 硬化 → 木化 ※ 髄 pith: 茎(幹)・根中央の基本組織で主に柔組織からなる (→ 放射髄 ray pith) = (樹木では)樹芯 → 心目: 髄を中心とした半径3 cm位の部分 [ 内側 ]維管束分化 differentiation of vascular bundle形態的分化の判断基準木部: 仮導管・導管の分化 仮導管細胞・導管細胞 - 二次壁リグニン化 → 成熟 = 原形質消失 師部: 篩細胞・師管の分化 (視覚による判断困難なこと多)
篩細胞・師管 - 細胞壁厚化開始 = 分化開始 核消失、カロースcallose形成 → 成熟 |
1922 Robbins, 1946 Loo, 1946 Ball: 茎頂切出し培養
原始的種 = 良く育つ + 大きく切出すとうまく行くが小さいとダメ ⇒ 分化決定時期特定 → intactな実験には限度がある1929 Piekington ハウチワマメ
1932 Helm: 双子葉植物で葉原基除去実験 → 葉跡分化
切除部分付近: 維管束なく柔組織ある - 求基的方向に除去の影響減る
→ Agar, sugar, inorganic salts中培養 1954 Wetmore: マツバラン・ハシドイ・クジャクシダ・トクサ・ミズヒキ・イソヒバ 200-250 μm3茎頂 → 小さな成体: 茎頂は全て分化能力持つ 1960 Ball: ソラマメ、アスパラガス茎頂培養系統調べる ⇒ 分化過程で細胞が相互の位置をずらす 0
P1: 未決定か決定 (canal可能) (< 30 μm l)
P13/P3 = 2/3がshoot, P10/P3 = 2/3がleaf, P14では影響ない 網状中心柱が管状中心柱になる 1969 Hicks & Steeves: ヤマドリゼンマイ 決定因子に頂芽関与
|
|
Def. 茎・根で木部-師部間に存在し分裂活動行う輪状細胞列の層 = 維管束形成層: 木部・師部形成のため分裂する細胞層 (= 分裂組織) (コルク形成層と明瞭に区別する際に用いる用語) 草本と木本草本: 形成層が分化しても大きく活動しない間に枯れる →
形成層から作られる部分は少ない ⇒ 木本性・草本性は形成層活動の連続性で決まる
= 形成層の活動は連続的な変化なので厳密には区別できない 形成層は茎だけでなく根でも分化する 発生: 一次木部と師部の間に分化が始まる →
一次維管束間の柔組織細胞が形成層化 → いずれも内側に二次木部、外側に二次師部を作る 側方分裂組織 lateral meristem: 肥大成長に関与 ☛ 木本植物茎肥大 → 中心柱外側の皮層・表皮部分 = 内部成長に遅れる → 死亡
(多くが)茎外側細胞一部が細胞分裂能復元 → コルク形成層分化
更に内側部分の一部が順次にコルク形成層化 - 年数経た樹木では元々の表皮・皮層消失 形成層起源維管束内形成層の起源前形成層 → 一次木部・一次師部 → 茎(普通は): 木部内側 - 師部外側 1966 原・駒嶺: ハッショウマメ黄化上胚軸 前形成層活動と形成層活動は不連続 ↔ 光照射個体では連続的変化 1965 Esau タバコ. 1967 Cumbie Canavalia:前形成層・形成層中間段階観察 - 連続的変化 1972 蘇: イチョウ - 様々な中間段階
⇒ 木本植物では一般に前形成層・形成層変化連続的 (それはそうだ) 形成層帯 cambial zone: 形成層と見做される一帯 ≈ 形成層 (細分する人もいる + 異常報告も多) 形成層タイプ一次維管束が密に並ぶため維管束間の柔組織部分が極めて少ない→ 維管束間形成層部分が極めて少ない Ex. ボダイジュ、タバコ |
一次維管束が適当な間隔置き分布
+ 維管束間形成層が作る二次組織は放射組織(柔組織) Ex. ヤマブドウ + 形成層から維管束内・間形成層の区別なく二次組織形成 細胞: 形成層構成細胞には2タイプ - 形成層の特徴的構造紡錘型始原細胞 fusiform initials: 細長い紡錘形
形成層内側 → 導管・仮道管・木部組織等形成 階層形成層: 接線縦断面で横に高さ揃った紡錘型始原細胞が並び階層状となる
系統的に進む双子葉植物 Ex. ニセアカシア、カキ 非階層形成層: 始原細胞配列不規則 - 隣接細胞間への割込成長
裸子植物に普通、原始的双子葉植物
放射組織形成 - 種により配列に特徴 1972 Carmi,Sachs, Fahn: 間隔は分化中の維管束組織が決める 割込成長: 形成層において →
紡錘型始原細胞増加 - 形成層細胞が形成層からはみ出す 二次維管束: 縦系と横系の組合せで組織構造決まる
縦系 = 紡錘型始原細胞由来
縦系: 裸子植物 = 仮導管主体、比較的一様 接線縦断面: 階層材 = 縦系要素配列が規則的 ↔ 非階層材 = 不規則 放射組織接線縦断面 → 単列放射組織 = 縦1列に数細胞並ぶ ↔ 多列放射組織 = 多数の列 二次師部: 二次木部の様に多年にわたり蓄積されることはない(1-2年多)コルク形成層を分化させる → 樹皮形成 組織培養による証明: オーキシンとサイトカイニンの濃度大きく変化カルス形成 → 組織分化 |
|
= 道管 + 仮道管組織 + 木部繊維組織 + 木部柔組織 (複組織) Def. 管状要素 trancheary elements = 導管 + 仮導管 (= 水を通す部分)
細胞壁肥厚が部分的に大きく異なる Ex. 環紋、螺旋紋、網紋、孔紋 導管(道管) vessel被子植物の木部主要構成要素 - 穿孔した導管要素が結合したものDef. 穿孔 perforation: 導管の孔 - 個々の細胞の上下にできる 導管は細胞が縦に並び管状となる - 仮導管より水移動性高い 水分通導作用c水が実際に通るのは主に導管・仮導管
大きさ・配列状態が種による特徴 → 種識別(系統関係)の手掛り 導管: 隣接細胞間で穿孔が貫通する ⇔ 仮導管: しない一次木部 primary xylem: 前形成層から分化する木部
頂端分裂組織由来 = 原生木部 + 後生木部
内原型 endarch: 前形成層構成細胞の茎の内側から分化 ↔
階紋導管 scalariform vessel: 梯子状に2次壁による肥厚が起こる
= 維管束形成層の活動で内側に作られた細胞から分化 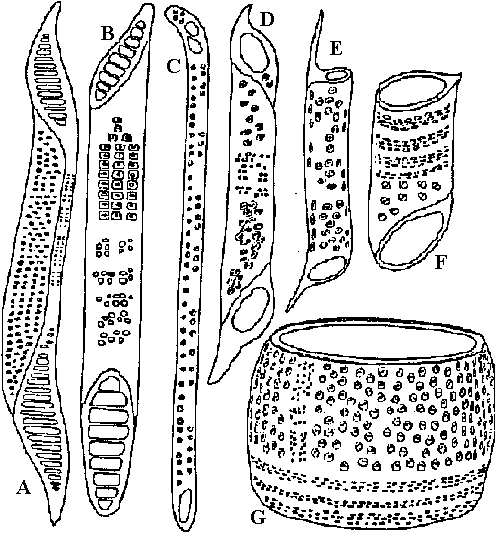 図32. 導管要素。A: Betula alba, B: Liriodendron, C: Lobelia cardinalis, D, G: Quercus alba, E: Malus, F: Acer negundo (Eames & MacDaniels 1947).靱革繊維: 茎皮部分にある丈夫な繊維。製紙、ロープ、織物等に利用 = アサ、コウゾ、Edgeworthia chrysantha
図32. 導管要素。A: Betula alba, B: Liriodendron, C: Lobelia cardinalis, D, G: Quercus alba, E: Malus, F: Acer negundo (Eames & MacDaniels 1947).靱革繊維: 茎皮部分にある丈夫な繊維。製紙、ロープ、織物等に利用 = アサ、コウゾ、Edgeworthia chrysantha羊歯植物木部: 仮道管のみで導管持たない(過去の認識) 一部の羊歯の根・地下茎で導管発見 (Carlquist & Schneider 2001) 管状要素の進化 (Carlquist 1961): 個体発生 → 系統発生仮導管 → [特殊化] → 導管 |
マオウ = 導管もつ → 裸子植物では派生的 双子葉植物: 導管は二次木部のみで形成 - 一次木部早期に形成開始 後者がより進化的 穿孔: 階紋穿孔(原始的) → 網紋・単穿孔 樹脂道 resin canal: 樹脂細胞から分泌された樹脂満たす管状組織
主に木部 (木部・師部両方に存在) 幹枝の長さに沿い配置される 水平樹脂道: 木部の放射方向に広がる樹脂道 木部放射組織(木部放射柔組織)と関連 粘液道 mucilage canal: 粘液を分泌・蓄積
水分保持、種子発芽促進 仮導管(仮道管) tracheid殆どの維管束植物木部で観察される組織
羊歯類・裸子植物の主要通導組織 壁孔 pit: 細胞壁にある小孔 - 隣接細胞間で液体交換 太さ0.005-0.060 mm、長さ1-6 mm
針葉樹: 体積の約90%以上占める 木部繊維細胞 fibrous tissue繊維細胞: 樹体支持作用受け持つ縦軸方向に細長い両端尖る細胞(1-2 mm)
繊維: 水の流れには働かない → vessel発達した被子植物にみられる(仮導管の退化したもの) 木部柔組織 xylem parenchyma木部唯一の生細胞 - 養分貯蔵(古語で細胞壁を細胞膜と呼ぶため膜孔と呼んだ) 壁孔(膜孔) pit二次肥厚した細胞壁のうち薄い部分。中層と一次壁のみからなる導管・仮道管で主に見られる 壁孔: 二次壁を持つ細胞同士間の連絡機構膜孔形成部に二次壁できない → 水移動部分となる 穿孔: 細胞壁に生じた大きな穴 - 導管
有縁壁孔 bordered pit: 二次壁が斜めに肥厚 二次壁の開孔部分直径が一次壁だけの部分の直径より狭い - 水流調節 互生壁孔 alternate pit: 単壁孔と有縁壁孔が交互に配置対生壁孔 half-bordered pit: 壁孔が規則正しく向かい合う 装飾壁孔 vestured pit: 壁孔の内側に小突起vestures存在
壁孔膜変形を抑え水流安定させる |
|
= 師管(被子植物多) + 篩細胞(羊歯・裸子植物) + 師部繊維 + 師部柔組織 (複組織): 能動的輸送(可能) ↔ 死細胞である木部は受動的水移動 光合成産物(炭水化物)を樹体の各部に輸送する組織 nutrient transportDef. 篩要素 sieve element = 師管 + 篩細胞 = 通導部分 篩要素分化過程: 核消失 + 液胞膜消失 + 液胞・細胞膜区別消失
+ 小胞体バラバラ ⇒ 原形質分離を起こす性質は消えていない 師部: 木部に比べ化石になりずらく研究遅れる (Esau 1970) Def. 師域 sieve area: 篩要素細胞表面に多数の篩孔集まる領域
= 師板のある範囲
= 篩要素細胞間物質輸送通路
傷を受けると速やかに大量に生産される (観察時に考慮必要)
→ 原形質消失 + カロース消失 (機能回復不可)
吻針を植物(師管)に刺し養分吸う昆虫 → 養分吸収中に麻酔 →
師管中圧力 = 30 atm |
Def. 篩板 sieve plate: 特定領域特殊化 = 師管細胞間にある板状の壁
→ 師管細胞間の連絡 1972 Satterthwait & Schope: ☛ Rhynia 篩細胞的な細胞 + 膜孔域様な部分 原生師部 protophloem → 後生師部 metaphloem (導管同様)
原生師部: やがて機能失い押し潰され見えなくなる 二次師部 secondary phloem: 形成層から分化 伴細胞 companion cell: 被子植物のみ - 発生由来は師管と同じ細胞 核がある - 核染色で普通の柔細胞より濃く染まる
液胞少、ミトコンドリア多、小胞体ある 師管付着柔細胞 parenchymatous cell - 師管原基細胞縦裂で形成 師部繊維組織 fibrous tissue = 師部繊維 + 師部柔組織師部繊維 phloem fiber: 種子植物幹茎師部外側にある繊維細胞。紡錘形多 師部柔組織 phloem parenchyma: 薄い一次細胞壁 ≠ 木質化 → 柔らかい |
|
Def. 維管束植物の茎や根の中心付近、維管束組織を含む領域 原定義: 内皮に囲まれた中の部分 - 内皮が多様なため上記定義 1886 van Tieghem: 茎・根組織系 = 表皮 + 皮層 + 中心柱 (用語登場)Def. 内皮 endodermis: 中心柱を囲む最内層の細胞層 根で明瞭に分化 ↔ 茎では様々(明瞭-不明瞭)、形態様々 A. 原生中心柱 protostele根・茎等の軸中心に木部位置し、その周囲に師部 Ex. ウラジロ、カニクサA0. 包囲維管束 concentric vascular bundle: 木部が師部を囲むよう配置 木部が外取巻く = 被子植物 ⇔ 師部取巻く = 羊歯植物(稀被子植物) A1. 単一中心柱(単純原生中心柱、原始中心柱、単中心柱) haplostele
多少なりとも円形の横断切片木部を持つ Ex. 水生被子植物: マツモ、スギナモ、フサモ A'. 放射中心柱 actinostele = 放射維管束 radical vascular bundle
⊃ 殆どの維管束植物根 (Ex. Psilotum) 水生根のように特殊化した例外がある Note: ジャガイモ塊茎(地下茎) vs サツマイモ塊根
芽
光照射
根
発芽 Ex. Lycopodium (ヒカゲノカズラ) 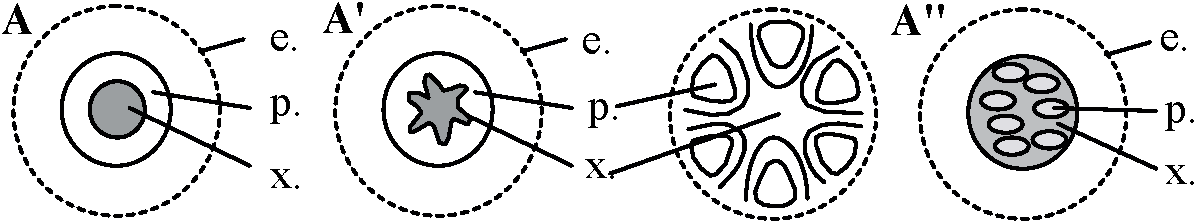
e: 内皮 endodermis, p: 師部 phloem, x: 木部 xylem 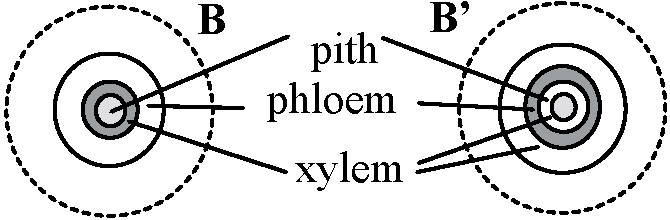
B. 管状中心柱 solenostele (s.l.) or siphonostele木部 = 管状 → その外面か内外両面に師部位置する Ex. フモトシダ
髄 pith: 維管束環より内側部分 B'. 両師管状中心柱 amphipholic siphonostele or solenostele, s.s. C. 網状中心柱 dictyosteleB'が葉隙に分断され、師部・内皮で囲まれた木部が横断面で不連続に見える |
Ex. 羊歯植物に多 (イヌワラビ、ホシダ) 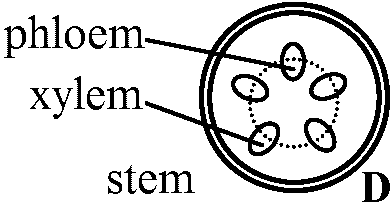
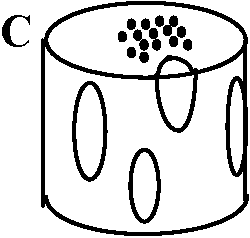
D. 真正中心柱 eustele= 並立維管束 collateral vascular bundle維管束が等間隔で環状に並ぶ - 主に双子葉植物茎
複並立維管束 bicollateral vascular bundle
双子葉植物 E. 多環中心柱 polycyclic stele木部か師部の一方が内外両側にあり他方を挟む = 2以上の同心円状の維管束組織を持つ中心柱
管状: 2環管状中心柱 dicyclic siphonostele、3環- tricyclic -, … Ex. 2環管状中心柱 = ワラビ 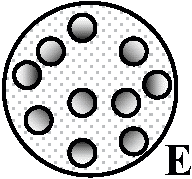
F. 不整中心柱(散在中心柱) atactostele= 外原型 exarch: 外端部分が成熟し、次に内側が成熟基本組織内にパターンのある分散をした維管束からなる
真正中心柱の一種としてもよい 中心柱説 stelar theory ☛ 進化1886 van Tieghem: 単一中心柱説 haplostele theory
仮定: 緑藻類 → 維管束植物祖先型出現 ⇒ 維管束単純
∴ Rhynia (化石種) = 原生中心柱が最も祖先的 基点: 原生中心柱 ⇒
→ 放射中心柱 1902 Brebner: 単一中心柱説の用語使用 ☛ 年輪解析 年輪 (year ring)樹幹横断面に同心円状に現れる模様中、毎年1輪ずつ形成されるもの= 形成層から作られた二次木部の1年毎の区切り 成長輪 growth ring ≈ 年輪 (※ 年輪は1年における1増分より大きいことも) 観察法: 切片染色(xylem, phloem分染) - 切片重ね合せ立体構造再構成シダではソーダで煮て柔らかくしたものを徐々に削り観察 |
[ 葉外部形態 ]
|
表皮系: 表裏各(多くは)1層(植物体保護)
= 表皮細胞 + 孔辺細胞 + 副細胞 + 毛, etc. 水・ガス通過しにくい構造。普通、表皮クチクラ層(外部クチクラ)の他に葉内部でも葉肉組織 mesophyll tissue や空気に接する細胞壁表面に薄いクチクラ(内部クチクラ)がある 鐘乳体: 表皮細胞外膜の一部が細胞内に向かい棒状に突出したもの
表面は炭酸石灰結晶が覆う Ex. Justicia procumbens, Ficus carica 2孔辺細胞が唇型に向かい合う構造 苔類除く全陸上植物胞子体世代に存在 気体(CO2, O2, H2O)が主に出入りする部分
多くの葉の表皮 → 向軸(表)側 < 背軸(裏)側 Ex. ムラサキツユクサ
☛ 気孔の開閉 柵状組織 palisade mesophyll + 海綿状組織: spongy mesophyll
陽葉 = 柵状組織発達 (↔ 陰葉 = あまり発達しない)
これらに加え厚角組織、異形細胞等
海綿状組織で富む - 柵状組織でも見られる
木部 = 導管(被子植物), = 仮導管(羊歯、裸子植物) vs 師部 = 師管 |
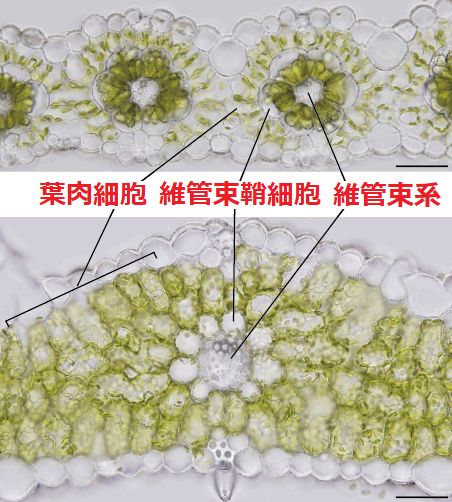
維管束鞘 bundle sheath: 維管束組織を取囲む細胞層
維管束鞘延長部 bundle sheathextension: 維管束鞘から表皮へ伸びる細胞列
C4植物 (シコクビエ) 葉の裏表(内部形態) ☛ 外部形態二面(背腹性)葉 bifacial (dorsiventral) leaf: 表面に柵状組織、裏面に海綿状組織を有する葉両面葉 isobilateral (isolateral) leaf: 葉の両面に柵状組織を持つ葉 クランツ型 kranz typekranz (独) = 花冠、花環,リース → C4植物の葉身に特徴的細胞内構造が発達した維管束鞘細胞が維管束系を取り囲み、その外側を1層の葉肉細胞が放射状に取り囲む 維管束鞘細胞: 葉緑体,ミトコンドリア、ペルオキシソーム多数存在
非クランツ型: 複数層の葉肉細胞が維管束鞘細胞取り囲み、維管束鞘細胞の細胞内構造は発達していない ≈ C3植物 葉組織間相互関係
1952 Wylie: 細胞-細胞接触面大 → 物質輸送大 |
|
Def. 発根 rooting: 根が成長する過程 (≈ 活着 rooting or establishment) 種子発芽時における根の発達
コムギ根: 発芽後40-50日で1 m lにまで成長 – 根先端分裂細胞 幼根 radicle: 種子中にある胚の幼い根 seminal root 胚軸下端にあり成長し主根 taproot or main rootとなる Def. 主根: 胚幼根由来の根 (裸子植物・双子葉植物) ⇒Def. 側根 lateral root: 主根から側方に伸びる根 (∴ 双子葉・裸子植物)
比較的乾燥、O2豊富 Def. 髭根 fibrous root (= 不定根 adventitious root): 幼根余り発達せず、主根は間もなく成長止まり、茎下部から多数根が出て細根rootlet束生し形成 主根なく、地上部大型化に適さない 構造: 水吸収に対し適応
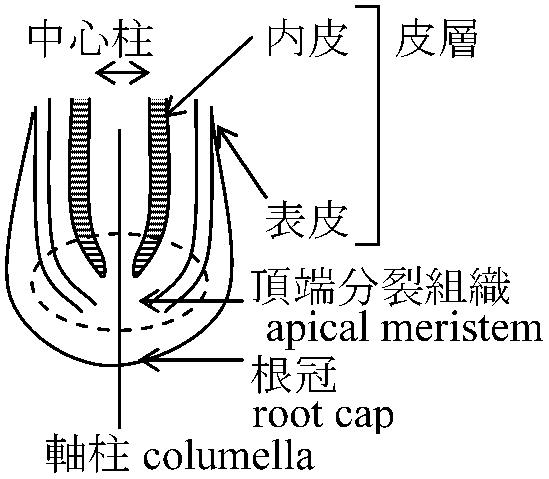
導管 xylem (死細胞) 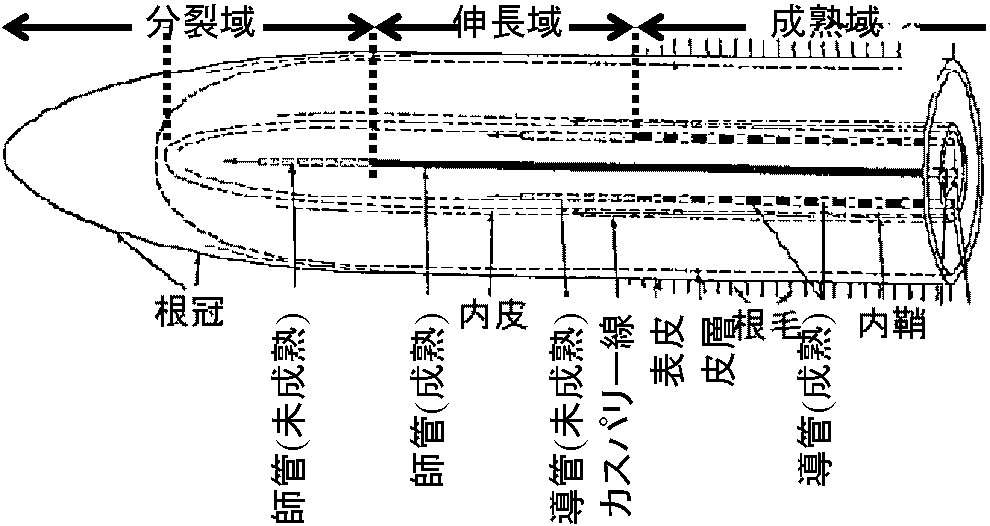 Def. 根端 root apex or tip = 根先端と近傍を漠然と指す (s.l.)
= 頂端分裂組織 + それに由来する一次組織 (s.s.) 環境に応じた可塑性高 Ex. 水耕すると根冠小さくなる
軸柱 columella: 根冠中央部分
中心柱(維管束と髄)・皮層・根冠分化 - 根端成長形態による分類試み 土壌: 大気と較べ安定環境 - 比較的原始的形質残る 根端分裂組織 root apical meristem根冠で保護される → 根冠・根表皮・皮層・中心柱を形成
発生・成長 = 伸長継続 ↔ 茎頂 = 周期性
貯蔵に使われる厚い層 Ex. サツマイモ |
側根: 根端から隔たった所に内生的に発生
根成長に伴い次の根毛形成 - 機能個所移動 内皮: 皮層の最内層とし分化 - 根では明瞭 物質は必ず内皮細胞中の原形質を通過(皮層細胞 ↔ 中心柱) カスパリー線: 内皮細胞細胞壁の特徴的構造 - 一次壁の一部 リグニンとスベリンの両方か片方を持つ 分裂域: 根端分裂組織ある = 細胞分裂盛ん = 細胞小伸長域: 細胞伸長盛ん(大きくもなる) + 細胞壁発達 + 形成層形成 形成層細胞 - 細胞分裂 → 根肥大 → 周皮形成 成熟域: 細胞分裂・伸長終わり = クチクラ層厚い
根端分裂組織タイプ (Guttenberg 1960, Popham 1966): 羊歯植物 = 1つの四面体始原細胞 → より高等な植物ではこのタイプは見ない 1917 Schüepp 本体冠体説 body-cap theory
本体・冠体で分裂様式異 = 機能異 その部分の全てがT/逆T状に分裂するとは限らない コルメラ columella: 横分裂を繰り返すことで形成される部分 1961(総説), 1967 Clows 静止中心説 quiescent center theory1952 Buvat: 待機分裂組織説: 茎頂頂端分裂組織中央に細胞分裂行わない部分 - 静止中心 静止中心 quiescent center: 根端分裂組織にも - 根発達につれ形成 周囲幹細胞にシグナル送る - 周囲細胞の分裂促進 茎-根関係根: 放射中心柱 ↔ 茎: 様々(真正中心柱、不整中心柱等)胚軸部分で入れ替わり ☛ 挿穂 発根促進剤 root stimulator活力剤型メネデール(二価鉄イオン) ホルモン剤型: 活着改善(可食植物に非推奨多)
ルートン(ナフチルアセトアミド)
成分: 0.40% IBA (有効期限3年) クロネクス clonex 使用法:粉剤: 挿穂の切口にまぶしつける 液剤: 処理濃度まで希釈し挿穂の切口を6-24時間浸漬 |
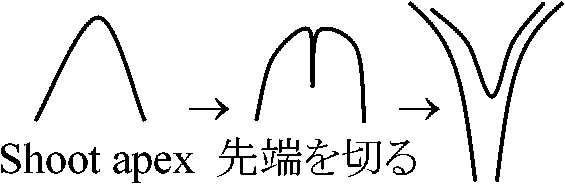 枝が2本(各々正常) → 予め決められていたら半分ずつの枝
枝が2本(各々正常) → 予め決められていたら半分ずつの枝