(2023年1月11日更新) [ 日本語 | English ]
HOME > 講義・実習・演習一覧 / 研究概要 > 小辞典 > 動物学 > 哺乳類
哺乳類分類階級表. 上位分類群の階級とそれに対応する日本語名称 (McKenna & Bell 1997, 哺乳類学会 2003)
巨目 magnorder Phylum Protochordata 原索動物門一生の中に (1) 脊索有、(2) 鰓穴持つ、(3) 囲鰓腔に囲まれた鰓腔持つ、(4) 内柱 endostyle 持つ、(5) 神経が時に神経管をつくる傾向 = 脊椎動物と共通部分あるClass Urochordata (尾索類) = Tunicata (海鞘綱) tunicates着生性/浮遊性Order Ascidiacea (ホヤ) 着生性。単性か群体。脳退化 Order Thaliacea (サルパ) 浮遊性。大部分が寒天質 Order Appendicularia (尾虫) 浮遊性。胴部・尾部区分明瞭 Class Cephalochordata (頭索類) = Leptocardia, lanceolets魚形、砂泥中棲息 |
Branchiostoma (= Amphioxus) (ナメクジウオ) 調査法個体追跡 individual traceテレメトリー(遠隔測定) telemetry: 行動圏を明らかにする 足跡: 行動特性、行動圏 糞・ペレット分析: 食性がわかる 食痕 トラップ
ベイトトラップ(餌罠) bait trap 足跡(足型) footprintロコモーション(移動能力) locomotion - 四足歩行 (歩行k ≠ 走行)趾行 digitigrade: 踵を浮かせた爪先立ちで歩行 Ex. イヌ科・ネコ科地上性哺乳類、全鳥類 蹄行 unguligrade: 踵を浮かせ蹄のみを地面につけ爪先立ちで歩行Ex. シカ、ヤギ、ウシ、ウマ、キリン、ゾウ等の蹄を持つ地上性動物 蹠行 plantigrade: 踵を含む足の裏全体を使って歩行Ex. 霊長類、クマ、パンダ、ウサギ、モグラ、ネズミ、コアラ |
Subclass Prototheria 原哺乳類亜綱 (原獣亜綱)= 原獣亜綱, 単孔亜綱Order Monotremata カモノハシ= 単孔類 (monotreme): 便等の出る穴が1つしかないことから命名卵生。子は母親から出る乳をなめとり育つ。哺乳綱中原始的で骨格も爬虫綱に似る Ex. カモノハシ platypus、ハリモグラ echidna (分布: オーストラリア-ニューギニア) Subclass Eutheria 真哺乳類亜綱 (真獣亜綱)Tribe Didelphia 双子宮類 (Metatheria 後獣類)胎盤発達していないOrder Marsupialia 有袋目(有袋類)骨格・繁殖法原始的 (オーストラリア)♀ 育児嚢pouchに嚢児産み暫く育児 → 子供は袋中の乳首に吸い付く Suborder Zoophaga 食肉
肉食 Suborder Phytophaga 食葉/草食
草食 Tribe Monodelphia 単子宮類= Placentalia 有胎盤類新生代に入り28(30)目におよび16目現存 Order Insectivora 食虫目真獣類中最も原始的 - 全真獣類はこれから分岐 多くは地下生活 (オーストラリア、南米中部-南部除く世界各地)口先長く突き出て鼻発達、目小。手足指5本 歯は門歯・犬歯・前臼歯・臼歯 Soricidae トガリネズミ shrew: 絶滅属 > 50 (始新世-現在) Talpidae モグラ: 「太陽に当っても他小型哺乳類と同程度に死なない」 ヒメネズミ*, Urotrichus ヒミズ*, Euroscaptor ミズラモグラ*, Mogera (アズマモグラ*, コウベモグラ*), カワネズミ, Neurotrichus アメリカヒミズ (N. gibbsii アメリカヒミズ, 北米) Order Tupaioidea (Scandentia) ツパイ= Tupaiidaeツパイ(キネズミ), tree shrews過去食虫目か霊長目に分類 小型で尖った吻と長尾 昼行性 昆虫や果実食 眼窩の外側が骨で囲まれる 手足指digitが良く働き、親指が4本の指と対向(霊長目の特徴) + 耳殻がヒトと良く似る Subfamily Tupaiinae ツパイ (5属17種)
Tupaia: コモンツパイ、ベランジェツパイ、ピグミーツパイ、ナガアシツパイ、ヤマツパイ、ニコバルツパイ、パインテッドツパイ、パラワンツパイ、アカオツパイ、ジャワツパイ、ホソツパイ Ptilocercus: ハネオツパイ Order Chiroptera 翼手コウモリ類 - 哺乳類中唯一空を飛ぶ形態: 親指除く手指が長く伸び、それらの指と後指間に皮膜発達 = 翼 親指小さく、翼前につく後足と尾間にも膜。大部分夜行性。耳発達し超音波発し反射音から周囲状況把握 腿間膜: 後足-尾間に発達している膜 – 一部の種 鼻葉: 鼻周囲に複雑な形の襞 – キクガシラコウモリ類やカグラコウモリで発達 生態: グアノ guano: 動物糞固まりできた堆積物 → 洞窟産グアノはコウモリ多 洞窟性動物 troglodyte: 主として洞窟で見られる動物
好洞窟性動物 troglophile: 一生を洞窟の内外どちらでも過ごす Suborder Megachiroptera 大翼手食植(果実食)性。視覚に頼り飛ぶ Pteropoidae オオコウモリ, pteropusオガサワラコウモリ、エラブオオコウモリ (ダイトウコウモリ、オリイコウモリ、ヤエヤマコウモリ)、オキナワコウモリ Suborder Microchiroptera 小翼手食虫性。超音波を出し、それを頼りに飛ぶHipposideridae カグラコウモリ Hipposideros turpis (イシガキ)コウモリ Rhinolophidae キクガシラコウモリRhinolophus (R. perditus ヤエヤマココウモリ) Molossidae オヒキコウモリVespertilionidae ヒナコウモリ モリアブラコウモリ*、モモジロコウモリ*、ヤエヤマユビナガコウモリ、フジホオヒゲコウモリ*、オゼホオヒゲコウモリ*、クロホオヒゲコウモリ*、クビワコウモリ*、コヤマコウモリ* Order Dermoptera 皮翼東南アジア - コルーコとか"飛ぶキツネザル"(ヒヨケザル類)Order Taeniodonta 紐歯目: 紐歯類!Order Tillodontia 裂歯目: 裂歯類!Order Edentata 貧歯目中央アメリカ、南アメリカアリクイ、地上性及び樹上性のナマケモノ、アルマジロ、グリプトドン類 無歯か、犬歯・門歯なくエナメルで覆われない歯 他哺乳目首骨は7個だが6-9個 Order Pholidota 有鱗目(センザンコウ類)東南アジア歯なく姿アルマジロに似るため貧歯目としたが体制異り別目とする 全身硬鱗に覆われ、毛僅かに鱗間に生えるか無毛 鱗は毛が集まり固まったもの 口先長く舌細長く前足は穴を掘る爪を持ちアリクイに似る ザンコウ等、長爪の動物はナックルウォークで歩行 Order Lagomorpha ウサギ (重歯)Order Cataceaクジラ (鯨)イルカ・クジラ類(哺乳類最大)一生水中生活 祖先は陸上生物だが面影ほとんどない |
Suborder Mysticeti ヒゲクジラ(鬚鯨)歯がなく「クジラヒゲ」を持つEubalaena セミクジラ black right whale, Balaenoptera ナガスクジラ finner whale, rorqual, Megaptera ザトウクジラ humpback whale, Eschrichtius コクジラ California grey whale Suborder Odontoceti ハクジラ(歯鯨)歯有 toothedクジラ whale: 大型, イルカ dolphin: 小型 (厳密な区別ではない) Supefam Physeteroidea マッコウクジラ: Physeter マッコウクジラ sperm whale Supefam Ziphioidea アカボウクジラ Ziphiidae アカボウクジラ Mesoplodon Gervais 1850 オウギハクジラ(扇歯鯨): M. stejnegeri True 1885 オウギハクジラ Superfam Platanistoidea インドカワイルカ Superfam Lipotoidea ヨウスコウカワイルカ Superfam Inoidea アマゾンカワイルカ Delphinoidea マイルカ Delphinidae マイルカ: Delphinus マイルカ common dolphin, Lagenorhynchus カマイルカ striped dolphin, Tursiops バンドウイルカ bottle-nosed dolphin, Globicephala ゴンドウクジラ piloy whale, Grampus (= Orcinus) サカマタ、シャチ killer whale Phocoenidae ネズミイルカ: Neomeris スナメリfinless porpoise Monodontidae イッカク: Monodon イッカク narwhal, Delphinapterus シロイルカ white whale Order Pinnipedia 鰭脚繁殖、休憩時陸上生活Phocidae アザラシ: 四肢歩行不可。耳殻なし(アザラシの子 pup) ゴマフアザラシ +、ゼニガタアザラシ +(南千島、北海道。唯一日本で繁殖)、クラカケアザラシ +、ワモンアザラシ +、アゴヒゲアザラシ + Otaridae アシカ: 四肢歩行。耳殻外向で見えるZalophus californianus japonicus ニホンアシカ*、カルフォルニアアシカ,ガラパゴスアシカ、トド+、オットセイ+ Odobenidae セイウチ: 四肢歩行1 gen. 1 spp. 2 ssp, Odobenus rosmarus セイウチ Order Hyracoidea イワダヌキ(岩狸)目= ハイラックス類 hyrax 1科3種 アフリカ-小アジア体は丸々とし四肢短く、形態モルモット様だが、臼歯はサイ・シマウマに近く、足は爪でなく蹄状 化石・遺伝子研究 → ゾウ等の原始的有蹄類と類縁関係 体温調節能低く、朝集団で体を密着させ日光浴後に採食
ハイラックス: 岩山、崖。イネ科植物食。大型。中近東-アフリカ全土 Order Proboscidea Illiger 1811 (長鼻目)化石始新世以降多量に産出 → 時代区分有効地上生動物最大(体重4-6 t)。骨組み頑丈 長鼻は鼻と上唇が、牙は上顎門歯が伸びたもの 暁象類: Archidiskodon ! Deinotherioidea 恐獣亜目Dienotherium! Elephantiformes ゾウ亜目Elephantoidea ゾウ 下目Elephantidae (ゾウ*), elephants* Loxodonta アフリカゾウ L. africana アフリカゾウ (syn. L. a. africana) L. cyclotis マルミミゾウ (syn. L. a. cyclotis) Elephas アジアゾウ/インドゾウ: E. maximus アジアゾウ E. m. indicus インドゾウ, E. m. maximus スリランカゾウ, E. m. sumatranus スマトラゾウ, E. m. borneensis ボルネオゾウ Order Sirenia 海牛目 (ジュゴン, sea cow, sirenian)2科4種: 形態アザラシに似るがゾウに近縁, 植物食 - 岸近くDugongidae ジュゴン (現生1種) Dugong dugon ジュゴン dugong, sea pig (北限奄美大島, 時に迷行)人魚伝説モチーフ Hydrodamalis gigas ステラーカイギュウ: 体長11 m、6 t wt。乱獲絶滅(1968) Trichechidae マナティー (現生3種) 肌色: マナティー manatee = 灰色 ↔ ジュゴン = 肌色-灰褐色 Order Primates 霊長目 primate☛ 北海道 Order Rodentia 齧歯 (ネズミ), rodentsLemmus Link 1795 (レミング/タビネズミ)Order Carnivora 食肉目 (ネコ)Order Tubulidentata 管歯ツチブタ類 = 奇蹄目に近縁な原始的有蹄類。欧州×・北米×・アフリカ普通1頭生活。前足でシロアリ巣を掘りシロアリ・昆虫捕食 穴を掘り3 m位のトンネルに巣を作る 有蹄類 ungulatas.l. 有蹄s.s. 真有蹄類 Euungulata (鯨蹄類 Cetungulata) =
ローラシア獣類 = 偶蹄目 + 奇蹄目 + 原蹄類 Protungulata (= 管歯目) Order Artiodactyla 偶蹄 (ウシ)Order Perissodactyla 奇蹄 (ウマ)奇数本(3か1)脚指 - 中指発達草食 → 消化法単純で、盲腸長く微生物を飼い反芻しない Suborder Ancylopoda 鉤足Chalicotheriidae カリコテリウム*Suborder Ceratomorpha 有角 (サイ)Superfam Tapiroidea バク(獏)上科 (1 gen. 4 spp.): 前脚4本, 後脚3本Tapirus バク Superfam Rhinocerotoidea サイAmynodontidae アミノドン: 絶滅, 雨竜炭田・宇部炭田・唐津炭田から化石 Hyracodontidae ヒラコドン: 絶滅 Rhinocerotidae サイ Subfam Indricotheriinae インドリコテリウム Suborder Hippomorpha ウマSuperfam Brontotherioidea 雷獣上科 (or Order Astrapotheria 雷獣目)南米 - 大型哺乳類(絶滅) Superfam Equoidea ウマ Equidae ウマ Equas ウマ (E. caballus ウマ), トカラウマ*, ノグニウマ*, ミヤコウマ*, ヤエヤマウマ*, ヨナグニウマ* ティタノテリウム |
[ 統合環境調査法実習 ]

食虫目 Insectivoraヒメトガリネズミ Sorex gracillimusオオアシトガリネズミ Sorex unguiculatus: 普通種 エゾトガリ S. caecutiens saevus: 普通種 トウキョウトガリネズミ S. minutissimus hawkeri 世界 world: トガリネズミ (Sorex) Long-tailed Ss. 欧・北米, ヒミズトガリネズミ(Blarinella) Asiatic Short-tailed S., コミミトガリネズミ(Cryptotis) Smallトガリネズミeared Ss., ブラリナトガリネズミ(Blarina) Shortトガリネズミtailed Ss. 北米, サバクトガリネズミ(Notiosorex) Gray Ss., or Desert S, オオサバクトガリネズミ (Megasorex) Giant Mexican S, ミズトガリネズミ (Neomys) Old World Water Ss. 欧・シベリア (Nesiotites) Mediterranean Ss., ケムリトガリネズミ (Soriculus) Asiatic Ss. 東南亜, ミズカキカワネズミ (Nectogale) Web-footed Water S., カワネズミ (Chimarrogale) Asiatic Water Ss. 日・ヒマラヤ, モグラジネズミ (Anourosorex) Mole S., or Sichuan Burrowing S. 亜, ジネズミ (Crocidura) White-toothed Ss. 欧・亜・Af, ザイールジャコウネズミ(Paracrocidura), ジャコウネズミ(Suncus) Musk Ss., or Pygmy Ss., house musk shrew 欧・亜, モリジャコウネズミ(Sylvisorex) Forest Musk Ss., ルウェンゾリジャコウネズミ(Ruwenzori sorex), オオアシジャコウネズミ(Feroculus) Kelaart's Long-clawed S., ピアソンオオアシジャコウネズミ(Solisorex) Pearson's Long-clawed S., or Mole S., モリジネズミ (Myosorex) Mouse Ss., or Forest Ss., クラカケジネズミ (Diplomesodon) Piebald Shrew, or Turkestan Desert S., コンゴヨロイジネズミ (Scutisorex) Armored S., or Hero S. 
翼手目 Chiropteraウサギコウモリ Plecotus auritus: 耳がでかい
齧歯目 Rodentiaネズミ Myomorpha生息地 = 森林性、草地性、住居性 森林性: 年繁殖回数地域差 - 本州低地: 春、秋2回。山岳地: 夏1回。九州: 秋-翌春1回 → 温度依存 ヤチネズミ Eothenomys andersoni (Thomas 1905): 東北地方以南 アカネズミ Apodemus Kaup 1829 アカネズミ A. speciosus Temminck 1844: 夜行性・単独性。主に森林に棲息(+ 河川敷や田畑)。低地-低山帯, 尾 ≈ 体長 → 形態的変異大 = 亜種 ホンドアカネズミ speciosus. サドアカネズミ sadoensis: 佐渡島. オオシマアカネズミ insperatus: 伊豆大島. ミヤケアカネズミ miyakensis: 三宅島 小型で毛色濃く腹部茶褐色. オキアカネズミ navigator: 隠岐諸島. ツシマアカネズミ tusimaensis: 対馬. セグロアカネズミ dorsalis: 屋久島. エゾアカネズミ Apodemus speciosus ainu (Thomas): 大きさ, 道 > 本州 ヒメネズミ Apodemus argenteus Temminck et Schlegel: 落葉層厚い安定した森林。木登り得意で半樹上生活。平地-高山帯。尾 > 体長ケナガネズミ Diplothrix legata Thomas 1906: 奄美大島・沖縄・徳之島固有 アマミトゲネズミ Tokudaia osimensis Abe 1933: 奄美大島固有 |
草地性 エゾヤチネズミ Clethrionomys rufocanus bedfordiae Thomas: 冬樹皮害(造林地)。開草地・湿地。尾 < 体長。(Irie & Tsuyuzaki 2007) ミカドネズミ Clethrionomys rutilus mikado Thomas: 尾短く木に登らない カヤネズミ Micromys minutus ハタネズミ Microtus montebelli: 平地-高山帯。草原的環境 (Ex. ハイマツ帯)。繁殖不定期、東北地方以南 住居性: イエネズミ house rat(俗) = ドブネズミ + クマネズミ + ハツカネズミ ドブネズミ Rattus norvegicus Berkenhaut: 移入、高湿度の所 クマネズミ Rattus rattus Linne: 天井裏など (アジア)ハツカネズミ Mus musculus Temminck et Schlegel: 移入  (エゾ)ナキウサギ Ochotona hyperborea (yesoensis)
(エゾ)ナキウサギ Ochotona hyperborea (yesoensis)エゾユキウサギ Lepus timidus ainu (Nomura & Tsuyuzaki 2015) アナウサギ Oryctolagus cuniculus: 移入、エゾユキウサギ競合(渡島大島) エゾリス Sciurus vulgaris orientis: 冬眠しない エゾシマリス Tamias sibiricus lineatus: 冬眠する エゾモモンガ Pteromys volans orii 食肉目 Carnivoraイイズナ Mustela nivalis (Linnaeus 1766)エゾオコジョ Mustela erminea orientalis (Ognev 1928): 20 cm ニホンイタチ Mustela itatsi (Temminck 1844) (M. sibirica itatsi) ミンク Mustela vison (Schreber 1777): 移入、エゾユキウサギと競合 エゾクロテン Martes zibellina brachyura ↔ ホンドテン Martes melampus: 道移入、競合 キタキツネ Vulpes vulpes schrencki 
 エゾタヌキ Nyctereutes procyonides albus アライグマ Procyon lotor: 移入、エゾタヌキと競合 (イヌ Canis lupus familiaris) (ネコ Felis silvestris catus) ヒグマ Ursus arctos 偶蹄目 Artiodactylaエゾシカ Cervus nippon yesoensis (C. nippon亜種: 道固有)頭胴長 140-190 cm、体重♂90-140(最大 200) kg、♀70-100 kg 洞爺湖中島個体群動態 (梶 1986) 1957 ♂1頭導入 + 1958 ♀1頭 + 1965 ♀(妊娠)1頭 20年間 - 指数関数的増加 1980 → 1981 → 1982 → 1983秋 → 1983-84冬153頭 ≈ 300頭(ピーク) 232頭(+ 95頭間引) 高茎草本消失 Deer line形成 ササ衰退枯死 剥皮増加枯死 蔓茎類・羊歯類減少 不嗜好植物増加 |
|
2科12属59種 楔状門歯持ち顎を左右に動かし採餌 (↔ 齧歯目: 下顎前後に動かし採餌) ウサギ Leporidae(rabbit, hare)Oryctolagus アナウサギ European rabbit日本産飼ウサギ Japanese White (白色種)の祖先、日本にいない)飼ウサギ: 体重4-6 kg。日本搾出唯一飼ウサギ品種。目赤。明治期に養兎流行時に輸入した外国種と在来種との間に出来たアルビノが起源? Lepus ノウサギ hare, jack rabbitL. brachyurus Temminck et Schlegel ノウサギ Japanese hare*本州、四国、九州 食痕: 鎌跡様な切口 糞pellet: 1 cm前後の押し潰された球形・褐色 食糞: 小粒状かクリーム状の糞(盲腸糞)を(即座に)食べる行動 → 盲腸糞は野外にまずない 夜行性 = 姿確認困難 → 足跡やフンで生息確認 地域的4亜種(冬季体毛白色度合いの違い): 夏毛冬毛 = 気温と日長で決まる
angustidens Hollister トウホクウサギ: 本州日本海側-中国地方山地。換毛し、冬毛は茶色型と白色型
ainu Barrett-Hamilton エゾユキウサギ(エゾノウサギ): 道全域 |
山地・草原・林。樹皮・小枝等主食 グリーンランド・アラスカ・北部カナダ Pentalagus ムカシウサギP. furnssi アマミノクロウサギ*奄美大島・徳之島(1963年特別天然記念物指定) 原始的タイプ: 目・耳小、足短い ↔ 爪はウサギでは長く1-2 cmあり頑丈 毛色、黒-グレー Romerolagus diazi Ferrari-Pérez (1893) メキシコウサギvolcano rabbitPronolagus アカウサギOchotonidae ナキウサギナキウサギ (周極要素、日本では北海道のみ)Ochotona ナキウサギ1 sp = エゾナキウサギ大雪山系岩石地帯。体重200-300 gと小型で、耳小さくネズミに似る。冬期も活動続け、岩場中に巣を作り、大量の餌を貯蔵。鳴声「キーキー」 |
猛獣。肉歯類(サーベルタイガー絶滅)。犬歯発達。臭覚・視覚優れるCanidae イヌ canine, dogCanis L. 1758 イヌC. familiaris イヌ (- domesticus ノイヌ)
野良犬(かつて飼われたイヌ(俗称)が危険 (純粋なノイヌはヒトから逃げる)
狂犬病発病時、脳神経節細胞内に生じる小体(出現率66-93%)
≈ 野犬: 狂犬病予防法等、公衆衛生分野使用)
非冬眠(冬も食料必要) + 群行動 1905 奈良県吉野郡東吉野村鷲家口で最後の捕獲記録 hattai Kishida 1931 エゾオオカミ*: 道 1900頃 最後の捕獲記録 Vulpes キツネ: V. vulpes schrencki キタキツネ*Nyctereutes タヌキ: N. procyonoides albus エゾタヌキ Felidae ネコ, catFelis ネコ族 little catsFelis catus ネコ cat+、Mayailurus iriomotensis イリオモテヤマネコ* (西表島), Felis (Prionailurus) bengalensis ベンガルヤマネコ Pantherini ヒョウ族 Acinonyx チーター族 cheetah Viverridae ジャコウネコジャコウネコ, ハクビシン+(白鼻心)Ursidae クマ, bear下臼歯3本Ursus arctos L. 1758 - arctos L. 1758 ユーラシアヒグマ, 基準亜種 |
- horribilis Ord 1815 ハイイログマ, grizzly bear - yesoensis ヒグマ+: 北海道
熊棚: 木に登り、枝搔き集め、実を食べ、残った枝を座布団とし使用
入る前に出没最新情報確認 → 見たらともかく遠ざかる 
 Selenarctos thibetanus japonicus ツキノワグマ (対策: ヒグマに準ずる)
本州(東北に大個体群)・四国・(九州絶滅) (環境省 2004) Ailuropoda melanoleuca (ジャイアント)パンダ Mustelidae イタチ, weasel下臼歯2本アナグマ badgar+, テン*(本・九・四・対馬), ニホンカワウソ* otter, ラッコ sea otter+ Mustela イタチ (M. erminea オコジョ, M. erminea orientalis エゾオコジョ, M. nivalis nivalis キタイイズナ(ネコ目最小), M. itatsi *ニホンイタチ, M. vison アメリカミンク) Martes zibellina brachyura エゾクロテン+ (道・樺太・シベリア・北欧) Procyonidae アライグマMephitidae スカンクHyaenidaeハイエナ |
* 日本固有(現世), + 日本分布, × 絶滅
|
偶数本脚指の有蹄動物 - 歩行時、中指-薬指間に体重かかる
反芻 rumination: する = 有角種多 ↔ しない = 牙発達 Bovidae ウシTragelaphus ブッシュバックBoselaphus ニルガイBos bosKobus ウォーターバックAepyceros インパラAntilope ブラックバックAntidorcas スプリングボックCapricornis カモシカ serow+♂♁角生え変わらない ⇔ シカ = ♂角皮袋形成し生え変わるC. crispus ニホンジカ Capra ヤギ goatOvis ヒツジ sheepOvibos ジャコウウシ(麝香牛) muskox, 1種O. moschatus Zimmermann 1780 (ジャコウウシ); 生息地: ツンドラ tundra 自生: カナダ北部、グリーンランド → 移入: Alaska, Sweden, Norway, ロシア北部等 食性: 植物食(草、ヤナギ等葉、果実、苔、地衣等)Cervidae シカ deerGiraffa camelopardali キリン(麒麟), giraffe(Giraffidae 独立見解)Alces alces, elkCervus L. 1758 シカ*現生4種C. albirostris Przewalski 1883 クチジロジカ C. elaphus L. 1758, アカシカ red deer C. canadensis Erxleben 1777 アメリカアカシカ elk C. nippon Temminck 1838 ニホンジカ Sika deer or Japanese deer 中国 - ロシア- 朝鮮 - 日本 (縄文遺跡 骨出土) 個体数爆発的増加(環境省 2013)
2011本州以南 = 推定261万頭 (2025に2倍), cf. イノシシ 88万頭 タンニン多い葉好む = イネ科草本、木葉、堅果、ササ(安定した重要食物) ♂♀別集団形成 → 交尾期(秋)♂♀集団形成 → 5-6月頃1頭生む ♂集団解散 + ♀集団に子供属す(普通、母子数頭群生活) ♂単独か♂同士小群 ♂泥遊びを♀よりする – 寄生虫よけ?(イノシシも) |
フィールドサイン: 足跡(2つ並んだ蹄)、フン(足跡追うと所々で確認)、食痕(両顎で引き千切る/樹皮剥ぎ)、角研ぎ 亜種複数見解: MtDNA分子系統解析 → 日本: 兵庫境に東西2系統分化 - yesoensis Heude 1884 エゾシカ: 道 (Tsuyuzaki & Takahashi 2007)
- yesoensis Heude 1884 エゾシカ: 道 (Tsuyuzaki & Takahashi 2007)
♂♀ 夏茶色全身鹿子模様。冬毛灰茶色無白斑
有珠でテントが囲まれた時は泣きそうだった...(ノイヌだったかも) - aplodontus Heude 1884 ホンシュウジカ North Honshu sika: 本州 - pulchellus Imaizumi 1970 ツシマジカ: 対馬
- keramae Kuroda 1924 ケラマジカ Kerama sika: 慶良間列島 (江戸時代に九州から移入) Dama dama ファロージカ (ダマジカ) fallow deer: 角先トナカイ角の様に先端部分が扁平し掌状 Rangifer tarandus L. トナカイ(馴鹿)reindeer, caribou♂♁共に角(♁ < ♂)。北極タイガ-ツンドラ Capreolus capreolus, roe deerMuntiacus reevesi, muntjac キョンSuidae イノシシSus L. イノシシ, wild boar or pigS. domesticus ブタS. scrofa イノシシ: ユーラシア大陸
本来昼行性 - 人為により夜行性となり農耕地等に出没 - riukiuanus リュウキュウイノシシ* Dichobunidae ディコブネEntelodontidae エンテロドンTayassuidae, peccary ペッカリーAnthracotheriidae アントラコテリウムHippopotamidae, hippopotamus カバOreodon オレオドン ×Camelidae, camel ラクダTragulidae, chevrotain マメジカ |
ヤク (yak)学名Bos grunniens L. (偶蹄目ウシ科)体長3 m程度(♂の方が大型)、体重500 kg程度、肩高2 m前後 高地適応。体表は蹄の辺りまで達する黒く長い毛に覆われる。換毛しない 肩がコブ状に盛り上がる ♂♁共に長さ1 mに達することもある長角を持つ 家畜としては、荷役用、毛皮用、乳用(乳製品含む)、食肉用に使われる
チベッタンの生活の殆どがヤクに依存している印象であった ロールガイ湿原 (Ruergai wetland) 1 1
 2 2
 3 3[1] ゾーネーションを調べた一サイト。白黒フィルムを持って行った(当時、論文は白黒写真が普通だったので) (Tsuyuzaki et al. 1990) [2] ヤク放牧地遠景。点々と見えるのがチベットの人たちのパオと呼ばれるテント。パオの周囲に沿って赤茶けた部分が裸地化の進んでいるところ。1988年7月21日。(Tsuyuzaki & Tsujii 1990) |
[3] 放牧地を近くで見ると、裸地化がよく分かる。中央で寝そべってるのとその他大勢が牛の仲間の1種であるヤク。結構危険! (1988年7月21日)
高尾山自然休養林の看板ホンドリス(ニホンリス) (リス科): 本州、四国、九州に分布するが、西日本に少ない。主にドングリやマツの実、クルミなど植物質のものを食べる。夏と冬で毛がかわり、冬毛には耳の先にふさ毛がある。イノシシ (イノシシ科): 夜行性で、円盤状の鼻で土を掘り返し、草や果実、小動物等を食べる雑食性。親子で生活し、子供は、はじめ体に縞がありウリの様なのでうり坊と呼ばれる。 |
|
類人猿 (anthropoid): ヒト以外のヒト上科(無尾) ≠ 生物学上の分類単位
Ex. ゴリラ・オラウータン・チンパンジー・テナガザル (Brace et al. 1965) 旧分類(一例)Order Primates 霊長(サル) primate全般的特徴: 新皮質発達 → 2つの目は並ぶが立体視可。色彩感覚有り↔ 臭覚退化 平爪(×ツパイ)、鎖骨良く発達。関節の可動範囲大きい一対の乳房持ち、普通一匹づつ子供を産む Suborder Prosimii 原猿/ニセザル表情筋未発達 + 鉤爪talon持つ種多歯櫛(下顎犬歯切歯鋭く細長く櫛の様に前に飛び出る)を持つ Infraorder Lemuriformes キツネザル SuperfamTupaioidea ツパイ(キネズミ/リスモドキ)
非サル的: 鉤爪 + 前顎骨 + 鼻粘膜常時湿 サル的: 眼窩外側を骨が囲む + 手足の少なくとも一方は親指が他指と対向 + 2つの目は並ぶが立体視可。色彩感覚有り SuperfamLemuroidea キツネザルSuperfamDaubentonioidea ユビザル Infraorder Lorisiformes ロリス Infraorder Trasiiformes メガネザル 原猿論: 原猿亜目 Ex. Tupaiiformes → 真猿亜目 Ex. Homo sapiens 雄と雌 / 夜行性か昼行性 霊長目の社会構造の原理
昼行性: 重層 (ペアタイプ単位集団 - 群タイプ単位集団) vs 単層 Suborder Anthropoidea (Simiiformes) 真猿/ヒトニザル表情筋発達(サルらしいサル) = 原猿以外メガネザル: 真猿類により近い → 分類群変更(原猿類廃止) Infraorder Platyrrhini 広鼻 Superfam Ceboidea オマキザル Infraorder Catarrhini 長鼻 Superfam Cercopithecoidea オナガザル Superfam Hominoidea ヒト: 尾なし 真猿論: 新世界猿社会 + オナガザル科群型社会 (群型社会通時的構造、群型社会発展、類人猿社会) 多くの種は樹上生活 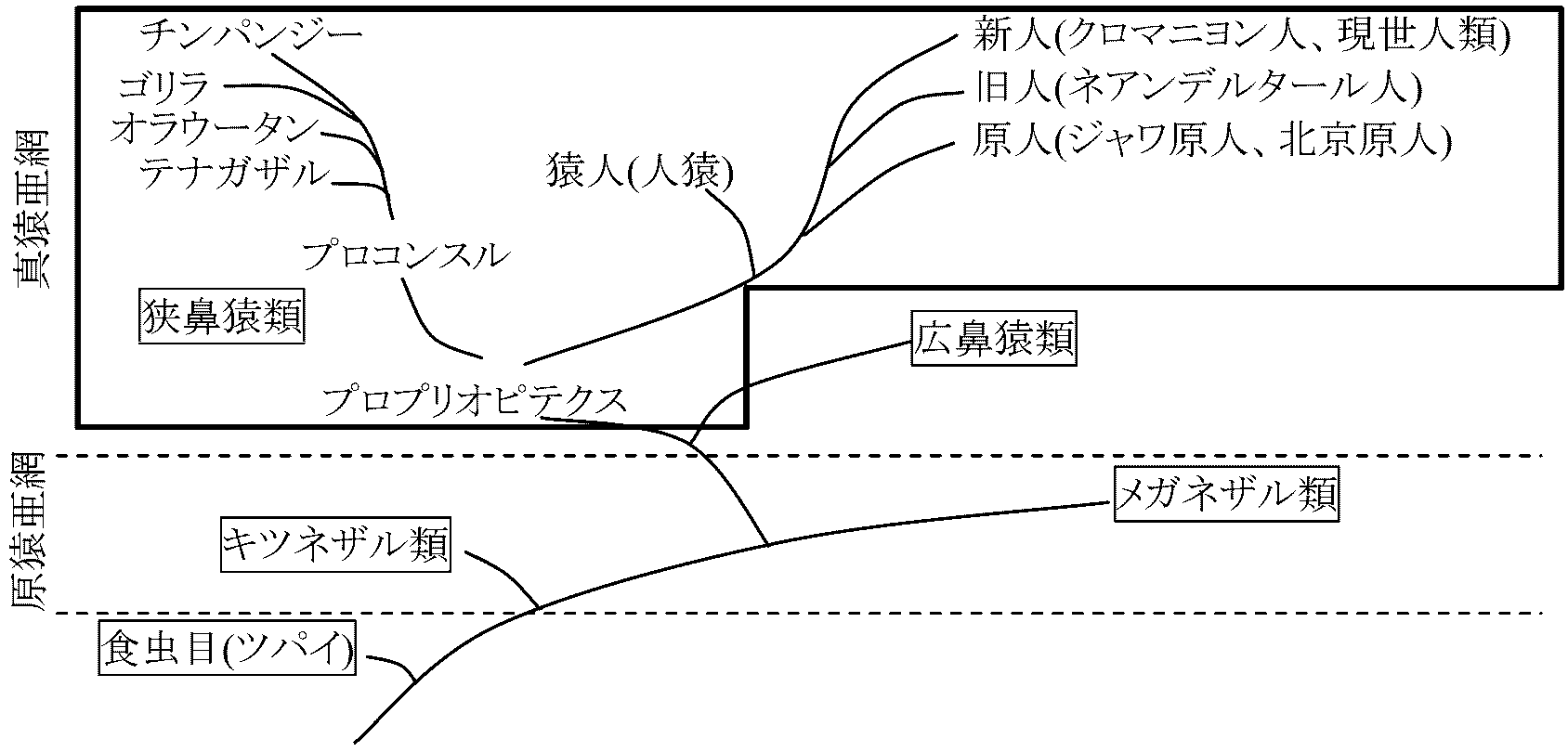
新分類(一例)Suborder Strepsirrhini 曲鼻(猿)鼻腔屈曲 → 鼻孔が左右に離れて外側を向く
タペータム(反射膜): 網膜裏側にあり弱光でも感知 → 多くが夜行性 Lorisidae ロリス(ノロマザル) Galagidae ガラゴ Infraorder Lemuriformes キツネザル型 全種がマダガスカル島および周辺に分布 Superfam Lemuroidea キツネザル Cheirogaleidae コビトキツネザル Indriidae インドリ Lemuridae キツネザル Lepilemuridae イタチキツネザル Superfam Daubentonioidea アイアイ キツネザル下目から独立 (1科1属1種) Daubentoniidae アイアイ Daubentonia madagascariensis Gmelin 1788 アイアイ: 1957年再発見 Suborder Haplorhini 直鼻(猿)= Simiformes 真猿類 + メガネザルInfraorder Tarsiiformes メガネザル型Tarsiidae メガネザルInfraorder Simiiformes 真猿型新世界サル new world monkey (中南米) Parvorder Platyrrhini 広鼻鼻形丸。尾活用Superfam Ceboidea オマキザル Atelidae クモザル Cebidae オマキザル Subfam Callitrichinae or Fam Callitrichidae マーモセット MarmosetとTamarin
Subfam Cebinae オマキザル Pitheciidae サキ 旧世界サル (アジア・アフリカ), 除ヒト Parvorder Strepsirrhini 狭鼻鼻切長。頬袋、尻ダコ持つSuperfam Cercopithecoidea オナガザル (1科のみ) 双隆線性 bilophodontism: 大臼歯の内側と外側の咬頭の間に2本の隆線
ヒト上科: 原則下顎大臼歯は5個の咬頭 Macaca マカク: M. fuscata ニホンザル Japanese Macaque, M. mulatta (以前 Macacus rhesus) アカゲザル Rhesus macaque (日本外来) Mandrillus sphinx マンドリル mandrill Superfam Hominoidea ヒト 尿酸オキシダーゼ活性なし - 分類再構成 Hylobatidae テナガザル gibbon インドシナ半島-マレー半島-スマトラ島-ボルネオ島 Hylobates テナガザル (1属見解) + Symphalangus フクロテナガザル (2属見解) + … (大型類人猿 = チンパンジー + オランウータン + ゴリラ) Hominidae Gray 1825 ヒト 現在: 現世2亜科4属 (従来: 現世1属1種 (二足動物 biped) = human) Subfam Ponginae オランウータン(ショウジョウ, 猩々) (Pongidae) Pongo オランウータン: P. pygmaeus オランウータン orang-utan Subfam Homininae ヒト Gorilla Geoffroy 1852 (ゴリラ, 大猩猩) Pan チンパンジー: P. troglodytes (Blumenbach, 1799) 野生チンパンジー wild chimpanzee: 系統的にヒトと最も近縁なものを研究しヒトとの比較 (→ 個体群内相互作用研究発展) Homo ヒト: H. sapiens sapiens直立二足歩行 + 遅速歩行・方向転換可 (手 ≠ 特殊化 ↔ 足 = 劇的変化) 大犬歯発達、大臼歯磨滅度の差が他の真猿類と異なる 大後頭孔: 頭骨から脊髄出る穴 向き: 直立二足歩行動物 = 真下 ⇔ 四足動物 = 斜め後ろ 骨盤骨
四足動物: 細長く平ら。脊柱-骨盤は水平で、後肢は骨盤に対し直角 |
ヒト 人の直系 主な退化・痕跡器官
原人 ┤ ホモ属 眼窩上突起・歯隙・
│ 母趾対向性・下位肋骨
猿人 ┤ 人類 切歯縫合・毛
チンパンジー ┤ ヒト亜科 長掌筋
オラウータン亜科 ┤ ヒト科 采上襞
テナガザル科 ┤ ヒト上科 第三転子・尻尾・瞬膜
旧世界猿 ┤ 狭鼻下目 小臼歯・鋤鼻器・盲腸
広鼻下目 ┤ 真鼻類 触毛・子宮中隔
メガネザル類 ┤ 直鼻亜目 茎状舌骨・鼻甲介・鼻鏡
曲鼻亜目 ┤ 霊長目
(他の)胎盤類 ┤ 真獣類 卵黄嚢
有袋類 ┤ 獣類 烏口突起
単孔類 ┤ 哺乳類 腓骨・座骨動脈・頭頂眼
双弓類 ┤ 羊膜類
両生類 ┤ 四足動物 エラ・鰓弓・心耳
魚類 ┤
図. ヒトの直系の分岐系統図と主な退化・痕跡器官
ヒトニザル = 新世界猿 + 旧世界猿 鼻鏡 rhinarium: 鼻孔周囲の毛がなく湿った皮膚部分 - 臭覚器官1. 脊椎動物としてのヒト (human as vertebrates)身体の各部分がいつできたか(組成より推定)a. 血液・体液、水との関わり体重に占める水 (%): ネズミ 60-70 カエル 77 カキ貝 83 クラゲ 98→ ヒト200万年、哺乳類6000万年 (Cf. リンガー液、生理的食塩水、羊水) 血液成分 ≈ 太古の海水 = 始生代に形成された? b. 背骨人: 頚椎7、 胸椎 12 、腰椎 5、 セン椎 5、 尾椎 3-5、計32-34個からなる石灰塩 70%、有機質繊維 30%。約200万年前のヒトの骨でも同様
ヒト ───────── 胎児 (1ヶ月) 脊索 ┐
哺乳類 ↓ │
爬虫類 2億年 軟骨 │系統進化説
両生類 3億年 ↓ │
硬骨魚類 3.5億年 デボン紀 ← ヒト骨の起源 化骨*│
軟骨魚類 4.5億年 オルドビス紀 ┘
原索動物 ナメクジウオ > ホヤ > 筆石 > ウニ > 軟体 > 擬軟体
底生 *: 硬骨になること
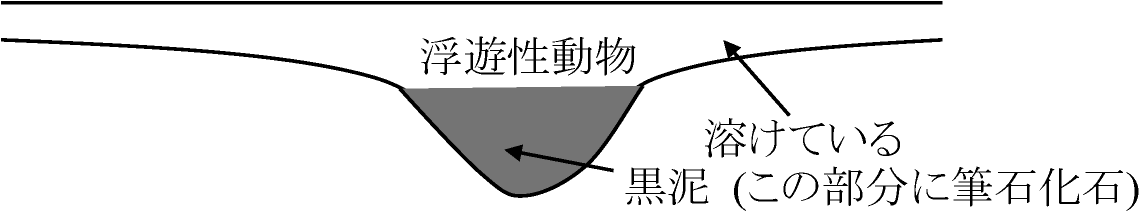
筆石: オルドビス紀示準化石(腸鰓動物。浮遊性)
二畳紀 ↑
石炭紀 ↑
デボン紀 ↑
シルル紀 硬骨魚 ↑ 内湾(泥の海)
オルドビス(筆石) ↑ ↑
カンブリア(三葉虫) 軟骨魚←浅海(海進): 無脊椎動物全ての門出揃う
__________✪_✪_✪_✪
原生代ヤツメウナギ: 現代形態的にもっとも下等な脊椎動物 ナメクジウオ: 脊索(原索)動物、浮遊性動物
カンブリア紀は新たなる動物発生の紀(例外ないほど浅海)であり原生代と明確な違いがでる。 それまでの動物は泳ぐこと、浮かぶことが不可能でありオルドビス紀以降の動物が出現する c. 四足・肺 (魚類 → 陸棲動物)哺乳類手足 - 魚類(肉鰓類)対鰭起源四肢動物の四肢の起源: 胸鰭 = 前肢 / 腹鰭 = 後肢 鰭の形態 鰓肺魚 無顎類 尾ビレのみ 板皮類 ┐ 鰭を完備 ⋄肺 – 交尾器(体内受精) =胎児栄養供給 軟骨類 ┘ 背骨。他の鰭出来る 硬骨類 = 水中を泳げる ⋄肺 ♥浮袋 両生類 四肢形成(デボン紀) 爬虫類 獣型(→ 哺乳類) vs 恐竜 2. 哺乳類としてのヒトa. 表皮毛皮 – ジュラ紀 / 毛 – 表皮の変形 - 陸・気候等により構成されたものb. 温血血球の小型化 – 体の隅々まで流れる。二心房二心室c. 歯の分化 (歯式 dental formula)ヒトニザル(2122/2122) × 2 or (2123/2123) × 2 (ヒト, オナガザル, ショウジョウ) + 顎・歯退化傾向 + 顎後退(下顎後方へ下がる) (2133)/(2133) × 2 オマキザル ニセザル
(2133)/(1133) × 2 メガネザル
(2123)/(1123) × 2 インドリ ウシ (0033)/(3133) × 2 = 32 イヌ (3142)/(3142) × 2 = 42 ブタ (3143)/(3143) × 2 = 44 (原始哺乳類 44) d. 有袋類白亜紀-現在 – 胎盤未完成(早産)。育嚢で育てる = 子宮類
歯数は一定しない。哺乳類と比べ脳頭部小さい e. 胎盤 ⇒ 胎生最古の哺乳類 – ゴビ砂漠で発見
│ 〽 ← 石灰質団塊(デルタテリジウム = 食虫類, 暁新世) 歯の生え方は我々の垂直交感 - 乳歯・永久歯(2生歯性) ゾウ: 水平交感 = 歯の方向の変化は鮮新世(100万年前)だが原因不明 f. 頭骨の変化↓ キツネザル・ 嗅覚 vs 視覚 = 双眼 + 色 (樹上)↓ メガネザル ↓ 温血でない種がある、夜行性 ↓ 有尾猿類 ↓ 下顎骨退化、四足(退化器官・痕跡器官vestige) ↓ 無尾猿類* 進化方向: 躯体大型化 – 肩・肘・胸発達 *: 中新世栄える g. 歩行 walkingRamapithecus → [直立歩行 upright walking] → Australopithecus[二足歩行 bipedalism] 1930 キース: 腕渡仮説 brachiation hypothesis
高等霊長類(四足歩行) → 類人猿(腕渡) → 直立歩行 腕渡 = 体幹直立姿勢前適応 + 大型化: 体重増加 → 腕渡 [反論] 骨・筋肉のつき方: ヒト ≠ 類人猿
Ex. 上肢: 腕渡をする類人猿 > ヒト 1979 Kondo (近藤): 小型類人猿仮説(モデル lesser ape model)
歩行様式多様化(= 特殊化していない)した中に二足歩行の起源 → ヒト: 特殊化 Ex. 土踏まず: 血管保護・体重移動 Ex.親指: 荷重に耐える Ex. 骨盤 pelvis: 漸進的進化 Ex. 下肢: 長い - 四足歩行困難 |
|
Def.(s.s.) 動物が摂取する食物の種類 (s.s.) Def.(s.l.) s.s.に加え摂食に関する習性や生態的特性を含む 雑食性 onivorous Ex. ヒト (雑食動物 omnivore) 肉食性 carnivorous Ex. バブーン (肉食動物 carnivore)
昆虫食性(食虫性) intestivorous
葉食性 folivorous: 葉 (食葉性害虫: 害虫の場合) 餌種数を基にした捕食者区分単食性 monophagous: 餌種1種 Ex. 一部寄生バチ狭食性 obigophagous: 分類上狭い範囲の数種 Ex. 多種寄生バチ 広食性(= 多食性) pobyphagous: 広分類群を餌種 Ex. ゴミムシ |
食性調査法ネズミ類: a. 食痕(種判定困難). b. 胃内容物鳥類: a. 目視観察. b. 胃内容物 + (糞 = 種子検出, Nishi & Tsuyuzaki 2014) 魚類・両生類・甲殻類: a. 胃内容物 摂食量(食害量) feeding rateDef. 単位時間当たりに動物が体内に取込む食物量 (Ex. kg/yr, kJ/yr) ⇒
摂食量 = 消化量 + 不消化排出量
野外測定困難 - 室内測定主 摂食様式デトリタスフィーダー: デトリタス(有機残渣) detritus を主に摂食する生物
腐食動物の特殊なもの |