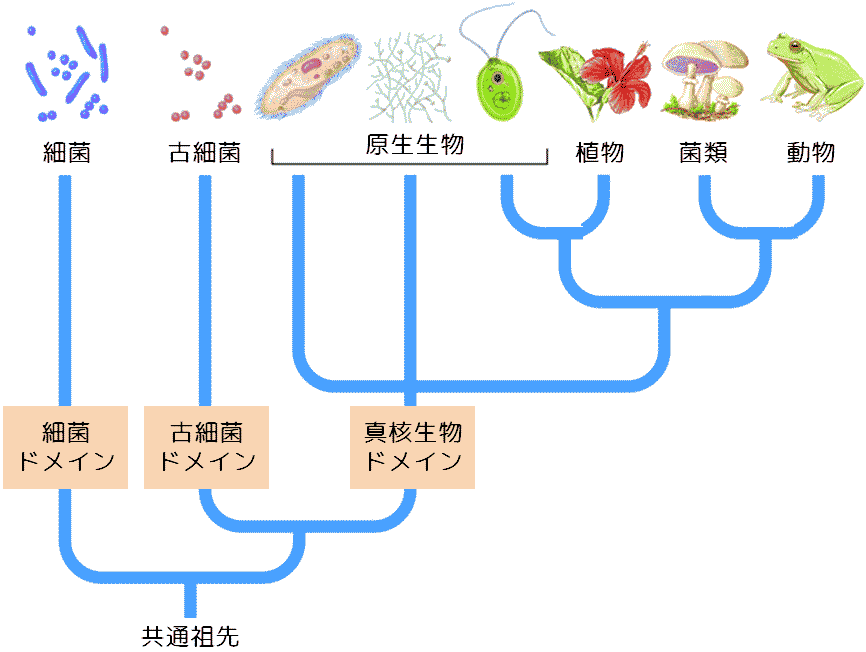
(2026年1月26日更新) [ 日本語 | English ]
HOME > 講義・実習・演習一覧 / 研究概要 > 小辞典 > 生物学
Def. 生命現象を様々なスケールで研究 (応用: 医学・農学等)
3つの生命概念
古典的分類動物学 zoology + 植物学 botany: 1980年当時の北大理学部生物学科教室区分 (生態学教室はない)⇓ 各生物群に関して 植物 (plant)
図. 5界説 動物 (animal)3界説 (three kingdoms)Haeckel提唱: 3 kingdoms = animal + plant + bacteria5界説 (five kingdom)1969 Whittaker RH提唱 → 植物動物に分けるのは便宜的なもの
Ex. 生物分類群: 完全な分類大系ない |
オーミクス(生物学)研究分野 = 研究対象 + omics (G. ome: 完全) ゲノミクス: ゲノム研究プロテオミクス: タンパク質の構造と機能について研究 全体主義と還元主義 (wholism and reductionism)個体以上の階層hierarchyを主に扱う各階層における相互作用 → 遺伝子レベルの手法が個体レベル研究に適用出来るとは限らない → 下の階層から上の階層は予測可能か 至近要因と究極要因 (proximate and ultimate factors)生態学研究目的 = [至近要因(近接要因) + 究極要因]解明 (Thomson 1950)= 生物現象の要因 = 発生機構(生理学的) + (進化的)存在理由 Ex. 鳥類の産卵時期の説明 (Baker 1938)
至近要因: 日長変化 → ホルモン分泌 → 生殖腺発達 相乗効果(シナジー synergy)要素が他要素と合わさる事により単体で得られる以上の効果を上げること1989 Covey SR "The 7 habits of highly effective people" (7つの習慣) Def. 相乗効果: 全体の合計が各部分の和よりも大きくなること Ex. 成長: 2植物を混植 > 個々に植栽 |
|
Aristoteles (Aristotle or Aristote) BC384-322 生物学の開祖: 動物・植物 (個体淘汰 = 適応) 無生物 → 下等植物 → 高等植物 → 海綿類・クラゲ類 → 貝類・昆虫類 → 甲殻類 → 頭足類 → 卵生類 → クジラ類 → 胎生4足類 → ヒト: ただし、種は不変(種の不変説) Plinius Maior (Gaius Plinius Secundus) BC39-79: 博物学・軍人・政治家
Naturalis Historia (博物誌) 全37巻
マルクス・アウレニウス帝の侍医 "De Rerum Natura (物の本質について)" → 動植物の起源述べる(進化) ↔ ローマそのものは神秘主義的思想 Botticelli, Sandro (1445?-1510): 「春」(1475)→ ルネッサンス期絵画の写実性 → 正確な観察への意欲 = 生物観察 Lamarck: 1808 「生物学 biology」の用語を用いるCuvier G 1769-1844, 仏: 古生物学樹立 – パリ郊外において脊椎動物調査
「地層中の化石を見ると下層のものほど現在の生物との差が大きい」 Scheuchzer, Johann Jakob (1672-1733), スイス、博物学(形態分類)
1709 洪水植物誌(Herbarium deluvianum): 植物化石図版化 Lyell, Charles 1797-1875 (Scotland, 地質学の父 the farther of geology) 1831 「地質学原理 Principles of Geology」(1830-31)
全ての地質現象や生命現象は過去も現在も同じ労力や経過で起こる → 一斉説: ゆっくりした過程 (進化 = 革命)。地球上の急激変化否定 Malthus TR 1766-1834: 人口論Jordan, Claude Thomas Alexis 1814-1897, 仏 1846 Observations sur plusieus plantes nouvelles, rares ou critiques de la France (rev. 1873) Viola, Draba材料に個体間差異記載 → 個体群間変異幅の差 = 地域差 → 地域個体群 local population = 微細種、小種 microspecies = ジョルダン種 Jordanian (Jordanon) – リンネ種で1種とした植物を20数種に細分: できるだけ純粋な小さい群に細分し、その各群を分類単位としての種の位置においた。遺伝学でいう純系に近い微細種 = 1種 → リンネ種の種内個々の変異型に種名を与える(cf. ダーウィン: 地域集団差に気づかない)
Jordanianは進化の基本であると考え、いち早くDarwinの考えを導いた 定向進化説 orthogenesis: 生物はその内的要因により常に一定方向に進化する(古生物学者ら支持) Ex. 馬化石、マンモス牙、アイルランドオオシカ巨大角 コープの法則(躯体大化の法則)
非特殊化の法則: 上位種族は必ず下位の非特殊化した種族から発生 Eimer T 1843-98, 独: 新ラマルク説についてアンモナイトを例に説明 Darwin Charles Robert: 自然淘汰説 Wallace AR 1823-1913, 英 博物学者 → 生物相互比較
1854-62 マライ諸島旅行: バリ島-ロンボク島間 = オーストラリア区-東洋区境界 (ウォーレス線) メンデル Mendel GJ: メンデルの法則 (Mendelian rules) ヘッケル (Haeckel, Ernst Heinrich 1834.2.16 - 1919.8.9 独, 動物学 1866: 動物形態学 (Generelle morphologie) (発生)反復説 (1872): 個体発生は系統発生を繰り返す = 進化論 + 分類学 個体発生: 個体発生過程 → 系統発生: その生物が辿ってきた進化過程 生物学大系化 → ecologyという用語提唱(1866 or 69)
Haeckelのecologyは適応生理学adaptive physiology分野 The nucleus = Natural history (博物学・自然誌・自然史のいずれかに訳される) → Taxonomy - Descriptive Schimper, K.F. 1803-1867 (Gr): Botanist (Schimper-Brawn's lawが有名)
生理主義 現在の生態地理学分野に属しSchimperの考えを生態学とし体系化 内容: life type分類 - 生物の獲得している生活様式 → 発展 = Plant ecology Wagner MF (1813-94)1868 隔離説 isolation theory: 各地を探検 → 種分化は地理的隔離による Jordan DS (1851-1931)
1872 Gulick, John Thomas (1832-1923): ハワイ諸島の巻貝
島ごとに異なる局限分布 → 局所的な地理的隔離も種分化に重要 Strasburger E (1844-1912)
1875 Zellbildung und Zellteilung - 細胞分裂観察 → 有糸分裂 |
Roux W Montogomery 1883 染色性 stainability のbody(後の染色体)の2分観察 Romanes GT (1848-94) 1885 生殖的隔離reproductive isolation:重視 →
生殖器官構造、生殖時期、性的本性変化等が種分化に重要 1885 定向進化説 (orthogenesis, determinate evolution) Waldayer WV 1888 W染色性のbodyに染色体chromosomeと命名 Henking N: 1891 相同染色体、X染色体発見 Montogomery: 1892 (相同染色体の)異親起源 Weismann A (1834-1914)
1893 ワイズマン生殖連続説(新ダーウィン説) 1895-: 有糸分裂mitosis、2n, n世代、減数分裂 de Vries H 1848-1935: オオマツヨイグサ1900 14年間8代自家受粉を行い子孫に1.56%の突然変異体観察
突然変異説 (1901): この突然変異は遺伝する → 突然変異 = 進化要因
2n = 14 → 2n = 28 (→ 27 → 26) Tschermak E (1871-1662, オーストリア): エンドウ ⇒ de Vries, Correns, Tschermak: 独立に遺伝研究 → メンデル論文発見 1900 [Mendelの法則再発見年] を出発点に遺伝研究発展 Sutton & Boveri: 1902-03 遺伝子染色体説 「遺伝子は染色体上に存在」 メンデル因子と減数分裂時の相同染色体の動き一致
体細胞 = 2n (2本づつ) → 生殖細胞 = n (1本づつ) Johansen Wilhelm (1857-1927) 1903 純系説 pure line (1909, 1911, 1913) Exp. インゲンマメ: 種子重による選抜 → 重い種子からは重い種子
数世代経過 - 同じ親株作った子株の平均種子重は親株と一致 ≡ 純系
変異を遺伝型変異と表現型変異の2つに区別
遺伝子型 genotype = 形成種(遺伝単位) - 遺伝する ⇔ 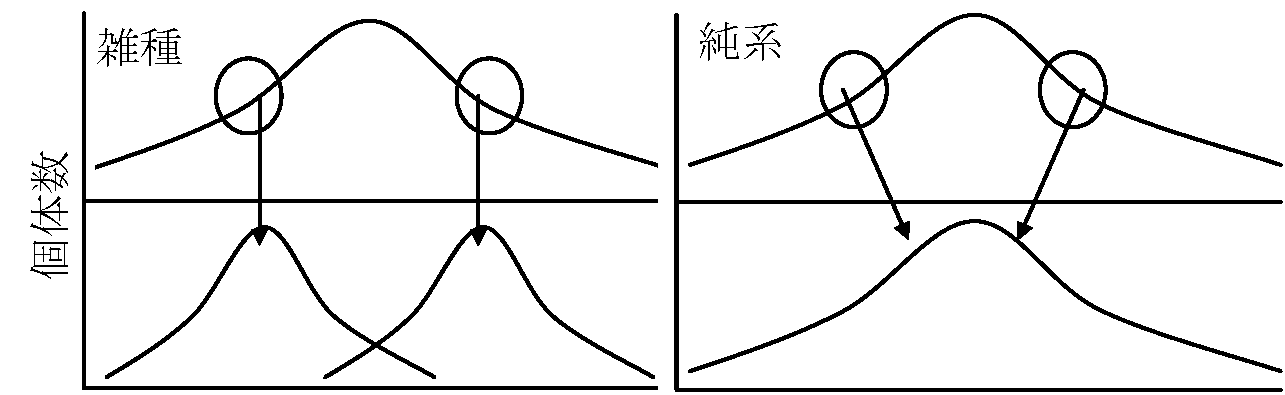 雑種時は淘汰有効 → 変異が一定方向に偏る ⇔ 純系 = どの個体を選んでも淘汰できない --- 1910年以前: 遺伝形質が染色体に載ることは否定的 ---Morgan TH 1866-1945, 米 1911, 1915 ショウジョウバエの遺伝に関する研究 ⇒ 1926 遺伝子説 連鎖linkage (cf. 交叉 corss over) → 遺伝子の染色体説確立 減数分裂における交換(部分交換)をもとに交叉率もとめ染色体地図完成 遺伝子説: 遺伝子は相同染色体の相対する位置に線上に配列
→ ペアンター、ドブジャンスキーら: 唾液腺染色体地図完成 = 核型としての形態的セット - 核型分析(LM levelで変異大) Vavilov NI (1887-1943): 古典的研究 (ルイセンコ論争で失脚 → 逮捕獄死)1926, 35, 51
1927 ショウジョウバエ突然変異誘導 (遺伝学, 優生学) X線照射 → 突然変異人為的に誘発 Dobzhansky T (1900-75): (生理的)隔離説1937 ショウジョウバエ集団遺伝学「遺伝学と種の起源」 1955 古典仮説classical hypothesisと平衡仮説balance hypothesis Dancer BH: Turessonの植物で提唱された概念を動物に導入 1929 Genetica 11: 399-450
Konvivium (F1稔性有) < Kommiskuum (F1不稔 = variety/race levels) < Komparium (F1兆候有 genus order) オザーク地方(Ozarks)全域のClematis個体群の分布調査 集 aggregate < 群 grade < 団 cluster < 帯 region < 界 range Morgan: 1960 Chromosomal theory of inheritance
遺伝子は染色体上にあることも証明 → 細胞遺伝学 (細胞学 + 遺伝学) 1965 科学的育種の基礎とし種・品種変異性に関する植物地理学的知識必要 |
|
Galenos: 生命 = 空気中にあるプネウマ(精のようなもの)が呼吸で体に入る
血液に取り込まれ生命の元になる 生気説 vs 機械説 (16世紀生命観)
Ex. 1. 有機物と無機物 2. 発酵と酵母 3. 形態形成 Goethe JWv 1749-1832, 独: 「若きウェルテルの悩み」
形態学研究も行い、変異を考察 生体を1個の全体として構成させるエンテレヒーという超自然的原理存在 1920- Bertalanffy, Ludwig von 1901-1972, オーストリア-カナダ
有機体論(生体論) = 2つの生命現象の特質 発生観 veiw of birth or genesis18世紀当時の対立仮説前成説 preformation theory (Cuvier 1769-1832): 卵精子中に個体の縮小物があり発生中大きくなる(宗教的伝説と重なり強い支持) ⇒ 両親に子供が似ることは説明できない – 否定
卵原説: 卵中に成体が入子に入る (精子発見まで主流) 蛹中に成体が折り畳まれて入っている → 卵に微小成体存在(卵原説) 1672 Reinier de Graaf (1641-1673) & Niels Steensen (1638-1686)卵胞発見 - 全ての動物に卵が存在する可能性 1677 Leeuwenhoek: 精子発見 (1675 Ham) → 精原説後成説 epigenesis theory: 卵は単純構造で発生過程で複雑な形が作られる(アリストテレス時代まで遡れる) Harvey (1578-1657): 1628 ヒトの血液循環証明
シカの腐った血から子供が生まれることを否定。卵生存在証明
→ ranular (= nucleus) → embryo formation Von Baer (1792-1876): dog oocyte確認 – 比較学手法の取り入れを主張
高等動物の胚は他の動物の胚と似ている Darwin (1809-1882): von Baer法則は進化論のもと説明可 (種の起源中) Müller JFT (1821-1897): Crastaceae研究からDarwinを支持 Häckel (1834-1919): von Baer法則は生物全体に当てはまる
Biogenetic law (recapitulation theory)「個体発生は系統発生の繰り返しである」 - 系統発生の短縮・簡略化はなされている
発生初期 vs 区別可能形質: 発生後期(= 進化後期段階) 発生学embryology: 観察と実験が絶対的なもの → 推論や理論だけではダメ Wilhelm Roux (1850-1924, 独): 2細胞期に割球を離す(= 発生生理学)
whole embryo – epigenesis ↔ partial embryo – preformation
2細胞を切り離すことは当時の技術で困難 – 片端を焼き殺す
2細胞ともに完全な胚となる = 前成説を否定[近代発生学出発点] 自然発生説論争天地創造時に使い残した想像力 → 自然発生 abiogenesis |
≈ 創造説 creationism 19c後半まで支配的 → 19c後半に発酵現象に生物を必要とするか否かの大論争起こる パスツール以前1668 Redi, Francesco 1626-1697, 伊: 腐肉を入れたビン口を布で覆う ハエ蛆は自然発生しない - 動物実験で自然発生説否定(不完全) 1675 Leeuwenhoek: 手製顕微鏡で微生物発見ジョブローの生物続生説(生物発生説) biogenesis theory 生物は生物のみから発生する 1749 Needham, John Turberville (1713-1781, 司祭, 英)加熱殺菌肉汁を熱灰中滅菌した薬瓶に入れ樹脂処理したコルク栓で密封 - 数日後、多数の顕微鏡的生物 → 自然発生説肯定 Spallanzani: 自然発生説否定1837 Schwann, Theodor (1810-1882, 仏): 加熱空気を煮沸スープに送る 滅菌空気 - 微生物発生しない 1854 Schröder: 殺菌綿を詰めた長い試験管 - 微生物発生しない1868 Liebig, 独: 発酵生物説批判 発酵後の不溶性物質(タンパク質)が酸化を受けその際にでた電子が糖に渡されて発酵が起こる 1859 Pouchet FA (1800-1872, 仏): "Hétérogénie"で自然発生説支持1962-1864 パスツール実験の欠陥指摘(ほぼ最後の自然発生説論争) 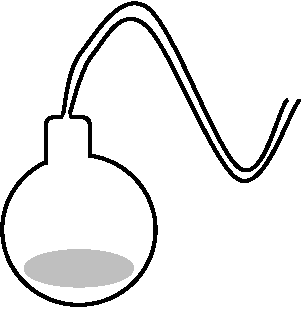 Pasteur, Louis 1822-1895、仏: 無発酵実験により自然発生説完全否定 (発酵研究にも大きく貢献)
Pasteur, Louis 1822-1895、仏: 無発酵実験により自然発生説完全否定 (発酵研究にも大きく貢献)1861 パスツールの実験 (微生物学の基礎) 1862 特殊フラスコ(パスツール管)使用し自然発生否定
生物は無生物からは発生しない → 親から子へ伝わる = 論争ほぼ決着(無生物から生物発生「全否定」ではなく、現地球条件下における生命自然発生を否定) 細胞説 (cell theory)細胞が生命の基本的単位である1590 ヤンセン親子, 蘭: 複式顕微鏡開発 → 物理学 → 微生物観察: 微生物 1610 Galilei: 顕微鏡作る 1665 Robert Hooke 1635-1703, 英: "ミクログラフィア"(1667) コルクガシ等観察 = 多数の小間仕切り cellula → 細胞cell発見 (観察は死細胞 → 細胞壁観察) 生理学 physiologyBell C 1778-1842, 英: 脊髄神経 Magendie 1783-1855, 仏: 脊髄神経 「実験生理学雑誌」創刊(1821) – 神経生理学確立 Bernard C 1813-1873, 仏: Magendie助手「実験医学序説」(1865) – 生理学全分野を扱う Müller JP 1801-1858, 独: 感覚研究門下にLudwig, Du Bois-Reymond, Helmholtz Ludwig CFW 1816-1895, 独: 物質交代。筋肉運動Du Bois-Reymond EH 1818-1896, 独 Helmholtzと電気生理学基礎確立 Helmholtz, Hermann Ludwig Ferdinand von 1821-1894, 独感覚生理学 – 視覚3原色説(1852) [物理: エネルギー保存の法則確立] 1831 Brown R 1773-1858, 英: 細胞に核発見 - 細胞説以前1838 Schleiden MJ, 独: 植物で細胞説提唱 1839 Schwann T, 独: 動物で細胞説提唱
「生物の体は全て細胞から作られ、細胞は構造上の基本単位である」 1846 Hugo von Mohl: 細胞の内容物を原形質と命名 1855 Rudolf Virchow: 「全ての細胞は細胞から」
細胞は存在する細胞から生じる – 無生物起源論否定
= 細胞は核のある原形質の固まりで、原形質こそ生命現象の基本単位 1875 Hertwig: 精核と卵核の合一観察 1879 Fleming: 有糸核分裂 1891 Driesch H 1867-1941, 独: ウニ卵の分離細胞が不完全だが胚に育つ 1924 Feulgen: 核内にDNA発見 1950 Andre Lwoff (1902- 仏, 1965ノーベル賞): 細胞説を言い換える 「生物世界を細胞レベルで観察すると同一性がある。どの細胞も原形質中に埋まる1個の核を持つ(基本構造一致)。代謝は本質的にどの細胞でも同じ(働き一致)。どの生物体も主要高分子を組立てる低分子は同じ(組立て一致)。生体系にみられる計り知れない多様性を作るために自然は厳選された素材を使っている。構造と機能の多様性、遺伝、種多様化の問題は僅かな種類の素材を使いこなし、特異的巨大分子ヘと組み立てることで解決されている - 各巨大分子はそれぞれ専門機能を負わされる。その役目を几帳面に果たすよう作られている。… 生体系がもしその勤めを果たさなかったら存在し得ないであろう。我々は、それがどの様にしてその任務を遂行するのかをありのままに学ばなければならない。」 |
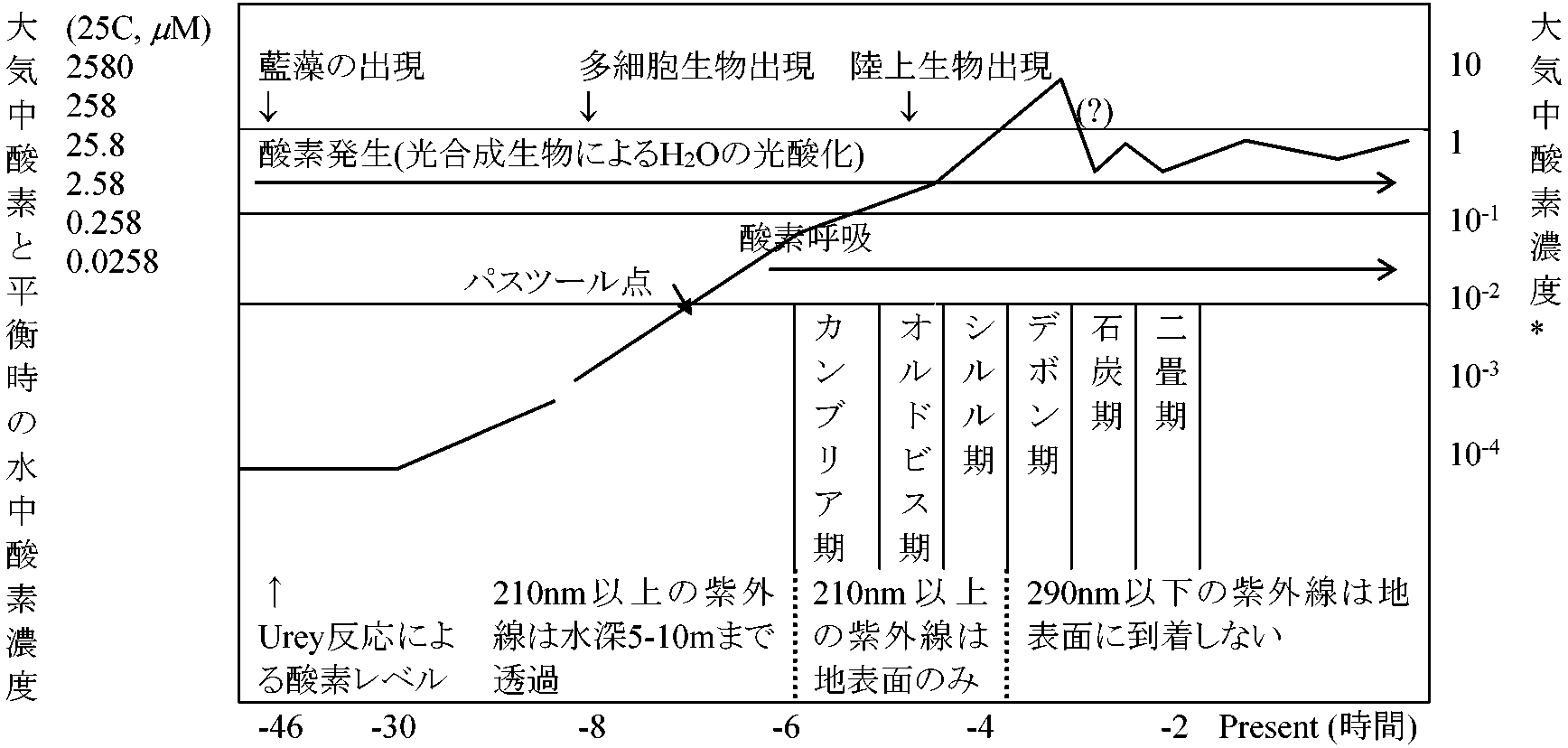
[ 初期生物: 化石による証拠 ]
研究法1. 人工生命構築2. 還元的アプローチ: ゲノム情報 → 原始生命推定
Ex. 現在の細胞が生存するのに必要な最小遺伝子数 = 推定250-300個 → 始原細胞(コモノート)推定
a) 生命構成全タンパク質はL型 生物学的分子進化1972: オーストラリアで隕石を落下直後回収(マーチソン隕石)
アミノ酸16種類 – 11種は殆ど地球にない。D/L型等量含 = 地球外合成 1871 Kelvin, Lord: 隕石説 - 隕石に生命(胚種)が付着し地球に到達 1908 Arrhenius S: パンスペルミア説(汎胚種説) Panspermia
径1 μm以下の微粒子は光圧を受け宇宙空間伝播可能 → 宇宙ダスト ≈ 乾燥バクテリアの赤外線スペクトル → 伝染病菌やウィルスは宇宙から飛来 1973 Crick & Orgel: ネオパンスペルミア説 neo-panspermia 地球外生命が生命を地球に送る 原始地球1) 原始大気: O2がなくH2の存在した還元状態
CH4, H2 (主成分) + H2O, N2, NH3, H2S, Ar等 2) 原始海洋 (35億年前): 火山活動による放出水蒸気凝結し原始海洋形成。大気中や地表の多くの物質は雨水に溶け原始海洋構成成分となる。海洋中は次第にCl-, SO42-, CO32-, Na+, K+, Mg2+等のイオンを多く含む 3) 原始地球での有機物合成のエネルギー源a) 太陽光線: オゾン層なく太陽光線の紫外線(UV)が強力に降り注ぐ。UVがH2O, NH3, CH4等を結び付け有機物合成に大きな役割を果たす b) 自然放電: 現在も雷放電が空中窒素固定に有効 (Miller 1950) c) 放射線: 原始地殻は今に比べ放射性元素多かった d) 火山活動 volcanic activities
熱水噴出孔: 原始のスープ形成 - 生命は深海熱水噴出孔で発生
高濃度 CH4, H2S, H2, NH3 + 金属イオン(Fe, Mn, Zn等) Ex. イアス地層(グリーンランド, 38億年): 海底起源玄武岩に硫化物堆積 修飾海水: Mo, Zn, Fe, Cu, Mn, Coイオン*濃度を103-5倍にした模擬海水
+ 9種のアミノ酸混合物 → 105°C (4週間) → 組織粒子生成 ⇒ 熱水噴出孔に見られる環境 *6種とも遷移金属元素 4) 有機物ができるまでa) UV(短波長光効果大)による有機物合成
原始地球: UV多量に注ぎ有機物合成
1952 Miller, Stanley (1930-2007, USA): 混合気体に放電 - 近自然条件条件でアミノ酸人工合成成功 1950' Fox SW, 原田馨: アンモニアとメタンを含む水蒸気を1000°Cに加熱 → 14種類のアミノ酸生成 5) 複雑な有機物合成化学進化: 前生物系 pre-biological system 成立までの過程
= 生命誕生まで(地球誕生から10億年後まで) C5H5N5 (アデニン) = HCN (シアン化水素)の5量体 1963 Ponnamperuma, Cyril (1923-1994), et al: メタン/アンモニア/水→ 電子線 → アデニン生成 (後にグアニンも生成確認) 小林憲正: メタン/窒素/水 → 火花放電 → 全核酸塩基生成確認原始細胞境界膜形成 → 自己組織化a) Oparin AI (オパーリン): コアセルベート説 coacervate theory 原始海洋にできた有機物が生命体になるには外界と境界でき、一定内部環境が形成される必要 → 境界膜としてコアセルベートの必要性重視 コアセルベート coacervates (CY): コロイド溶液中でコロイド分子が集合し濃厚なゾルとなり、外液から独立した顕微鏡レベルの小液滴 前生物系: プロトビオント protobionts: 原始海洋中でタンパク質は水を吸着してコロイド化し、次第に集合し大形粒子となり水の皮膜をつけた多分子系CYになって外部の海水から独立→ 何億年もの間に個々のCYは (1) 周囲物質を吸収変化させ成長、(2) 成長途中で2分し吸収力強め、(3) 他CYを取込み増大した |
前駆生命体 proto-organisms (原細胞 proto-cells): CYは長期間に様々な変化を遂げ触媒や有機物をも取込み最も優位な発達をしたものが成長・物質交代・分裂能力を持て、次第に原形質をもつ原始的生物に進化した b) Bernal JD (英, 物理学) 1951「生命の物的基礎」 - 生命の起源仮説提唱 低分子有機物 → 高分子有機物 → 原始生命形成
粘土: 生成物濃縮の際にの濃縮剤(吸着剤)として機能 核酸合成 = プロテノイドほど容易ではないが種々の条件下で生成成功 1961 Schramm G et al. (独)
ヌクレオチドとリン酸化合物混ぜ20-30個の核酸断片合成 i) Fox: アミノ酸乾燥混合物を数時間加熱脱水縮合反応
→ タンパク質様高分子重化合物(プロテノイドproteinoid)分子合成 プロテノイドミクロスフェア説: 原始細胞起源に関する一仮説 ii) 赤堀四郎(生化学者): ペプチド結合をしタンパク質合成 →
多量のエネルギー(ca 3 kcal/ペプチド1 mol)必要 6) 物質交代の進化と生物の発達 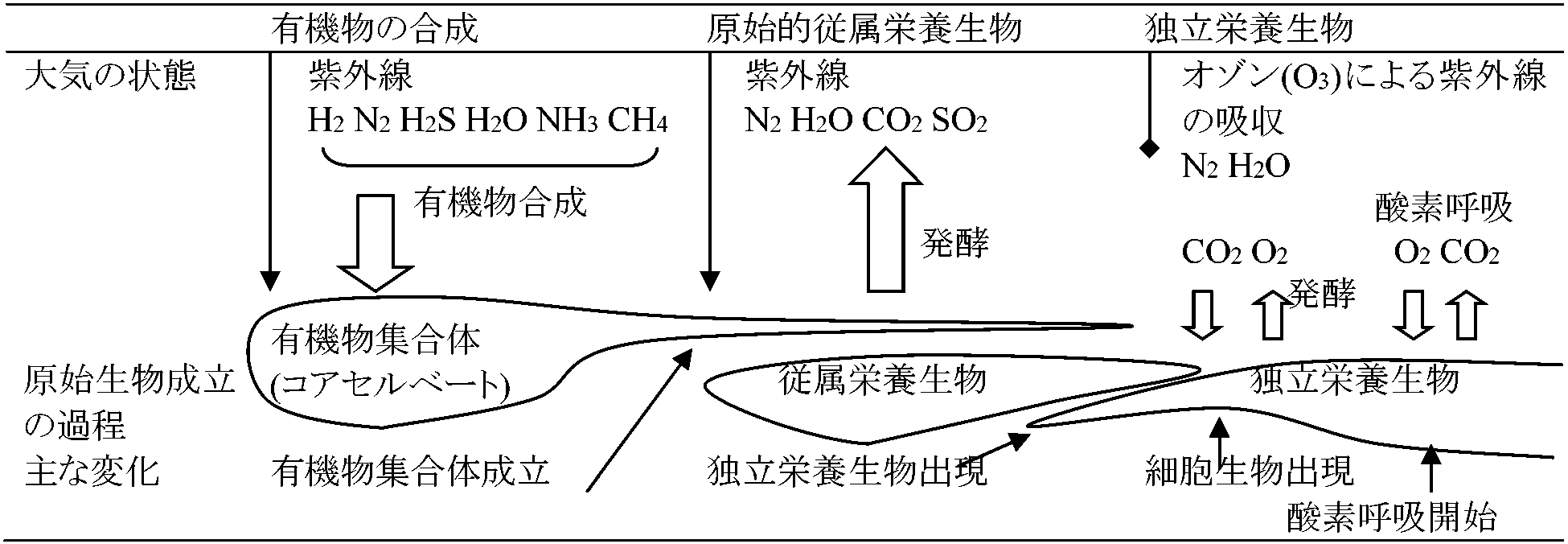 a) 独立栄養生物起源説(現在否定): 地球上で無機物から有機物合成できるのは炭素同化をする独立栄養生物のみ - 最初に生じた生物は独立栄養生物 b) 従属栄養生物起源説 (Oparin, Bernal) 有機物合成は生物存在しなくても起こる。最初に生じた生物は原始海洋に存在する有機物、物質を利用し生活する従属栄養生物 i) 最初の生物は原始海洋中に存在するアミノ酸や糖類を栄養分とし生活する単純構造従属栄養生物。原始生物は有機物をO2なしに分解しエネルギーを取り出し、現在の嫌気性細菌と似たもの。後に発酵機構が完成した酵母菌のような発酵生物が分化進化 ii) UV減少 = 気温↓(環境激変) ⇒ 海洋中有機物↓ + (無機呼吸により)大気中O2↑ ⇒ 原始的従属生物のエネルギー源↓ = 生存困難 ⇒ 大気中CO2やN2吸収し自己有機合成できる独立栄養生物出現 iii) 独立栄養生物も最初は化学合成細菌様であり、後に光エネルギー利用する光合成細菌様なものが現れ、次第にO2を放出する単細胞緑色植物へと進化した。そして光合成による放出O2量増加につれ、O2利用し高能率エネルギー獲得法である酸素呼吸機構が完成し、それと共に単細胞生物から多細胞群体を経て多細胞体の酸素呼吸生物が出現 大酸化イベント: 2度 2000 Farquhar J: 32S/33S比は通常大気では一定 ≥ 24.5億年 → 比が不安定 - 24.5億年以降に現在の大気に近づく 2008 加藤泰浩: 地下にある29-27.6億年前の火山灰に酸化痕跡29-27.6億年前の大気に酸素はあった - 同位体比もこれを支持 コアセルベート ⇒ 原始的発酵による従属栄養生物 ⇒ CO2, CH4増加 ⇒ ポルフィリン色素を持つ独立栄養生物 ⇒ 葉緑体を持つ光合成植物 ⇒ O2, CO2循環 ⇒ 呼吸作用発達(酸素呼吸生物出現) ⇒ 高能率エネルギー獲得 ⇒ 体制変化 [前核細胞 → 真核細胞、単細胞 → 群体 → 多細胞] ⇒ 多系統の生物へ進化
1981 Cech et al.: RNA - 触媒反応 ≡ リボザイム ribozyme 1986 Gilbert, Walter: 生命の起源 = RNAからなる自己複製系
生命は触媒機能と遺伝情報を兼ね備えたRNA分子から誕生 [問題] RNAの化学合成条件未発見 – RNA出現過程不明 |
生命科学 (life science)生命現象を明らかとしようとする学問領域生物学を中心に化学・物理学等の基礎科学と、医学・薬学・農学・工学・心理学等の応用科学とから総合的に研究 寿命 longevity一卵性双生児寿命は二卵性双生児より寿命一致率高い → 遺伝因子関与突然変異集積説 mutation-accumulation theory: 老化が体の構成細胞突然変異により起こる。体細胞突然変異集積により遺伝情報が誤って読み取られ、誤りを持つ酵素が細胞内に生じ、結局は細胞死滅、さらに器官活性低下等の老化現象発生 プログラム説 programmed aging theory: 1受精卵から出発し、ある細胞は肝細胞に、ある細胞は筋肉細胞に分化するよう、老化も分化過程同様プログラムされたもの。老化形質を発現する遺伝子(老化遺伝子/寿命遺伝子)は確認されている。老化開始させるスイッチがオンになると、遺伝情報、複製、転写、翻訳等の生命維持に不可欠な過程に誤りが生じ、細胞死、そして種々の老化現象を起こす 拮抗的多面発現説 antagonistic pleiotropy theory: 若い時に有利な遺伝子 → 後年に不利な作用 (進化的トレードオフ) 老化(加齢)老化(加齢) aging = 生物個体全体の老化過程(時間経過による機能低下)
老化作用 ageing action アポトーシス apoptosis自殺機構: DNA損傷 → その変異を伝えないよう自殺ネクローシス necrosis: 自発的ではなく環境に誘導される細胞死 短命植物 ephemerophyte: 短命な ephemeral (☛ spring ephemeral)Def. 適応 adaptation: 種がある環境で生存するのに有利な形質を持つこと 調節 adjustment 順化 acclimation (馴化 habituation) |
________[ ____ 精神 ____ ] 倫理はどうなる? - 優生学の復活?優生学 (優生思想) eugenics1869 Noyes, John Humphrey (1811-1886): ヒトによる優生学実験1883 Galtonが用いた: 良い種のギリシア語起源
遺伝的要因 = 人間の性質規定 → 因果関係利用したりそこに介入
強制的不妊手術(断種) + 施設隔離収容 等 1884 高橋義雄 「日本人種改良論」 - 黄白雑婚論 1910 海野幸徳 「日本人種改造論」 1930 日本民族衛生学会(日本優生学協会)設立 1932 Muller: 優生学学会で優生学に対する懐疑に関する講演 1948 優生保護法制定 現在: 選択的出生前淘汰の是非 + 遺伝子技術進展 = 優生思想否定
クローン人間(生物)の権利 - 年齢の異なる一卵性双生児 |
☛ 今西進化論(別格)
|
Def. 進化 evolution: 生物が世代を重ねる中で遺伝的形質が変化する現象 (ラマルク 1815-22) 用不用説 (use-and-disuse theory)キリン(ジラフ)の首 昔首短 → 高木の葉を食べ一生首を伸ばす → 親 < 子 < 孫と徐々に首が長くなり、何百何千代かけ現在のジラフになった= 自然発生を説き原始生命は進化の必然的傾向を持つ → 進化の副次的要因として用不用説を考えた
(ダーウィン 1859) 自然淘汰説 (natural selection theory)キリンの首 ジラフは昔首が短かったが、一群のうち首の長いものが生存競争に勝ち、その子の中でまた首のより長いものが生き残り現在のジラフとなるDef. 自然淘汰 natural selection: 同親から生まれた子間にも個体変異がある。生物は限られた資源に対し同種個体間競争が起こり(生存競争)、生存競争に有利な形質を持つものが生き残り(適者生存)、その形質は子孫に伝わる。このような関係が繰り返され(自然淘汰)現存する生物が生じた
突然変異と個体変異の区別をしていない (個体変異は遺伝しない)
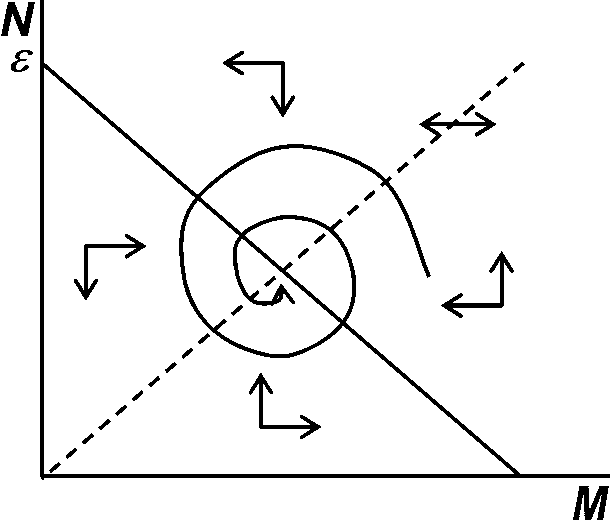
Humboldtの空間軸、Darwinの時間軸を合わせ2次元的解析導入によりEcology飛躍的に発展 淘汰万能主義1960s': 淘汰に関与しない中立的な対立遺伝子neutral allelesは存在しない現代進化説統合説 synthetic theory: 複数要因の総合された結果 = 進化= 突然変異 + 自然選択 + 隔離 進化 = 生物集団遺伝子構成の変化: 生物集団遺伝子構成が時間と共に変化する様式を調べる(集団遺伝学)。生物集団を個体群とみなさず、遺伝子集団としとらえ、その集団遺伝子構成がどの要因により、どう変化するかを調べ、法則性を見出し、進化のメカニズムを説明する
突然変異 ------------------------------------------------------→ 新種形成 関連進化理論1972 断続(平衡)説 punctuated (equilibrium) hypothesis
Eldredge, Niles (1943-), 古生物学, 米
進化の証拠1. 形態学 (解剖学): 生物形態・構造は環境で変化の方が説明しやすいa) 相同器官: 現在の器官構造は異なるが、元々発生の同じもの – 適応放散 適応放散 adaptive radiation: 同系統生物が異なる環境に適応した生理的、形態的性質へと変化していくこと
適応放散 ↗ ○ 適応集中 ○ ↘
Ex. 脊椎動物前足、哺乳類頚骨数はクジラもキリンも7。脊椎動物の浮袋、肺 Ex. 鳥類翼と昆虫羽、魚エラと貝類エラ、ブドウ巻きヒゲ(茎)とエンドウ巻きヒゲ(葉) b) 痕跡器官(退化器官): 祖先は使ったが現生物には痕跡的に残るEx. ヒト耳殻筋、クジラ後足 2. 発生学: 発生初期には、どの動物も胚形成や構造に差がない
発生途中で成体にない器官が一次的に生じる Ex. 哺乳類の鰓穴
i)_軟体動物 卵 → トロコフォア → ベリジャー → 成体 一時的に出現し消失する構造がある (ヘッケル: 反復発生説) 3) 生理学 a) 血清抗原抗体反応 |
Ex. ヒトと近縁種ほど血清中タンパク質に共通部分多 - 抗原抗体反応差小
ヒトの血清をウサギに注射 → 2-3週間後に再びヒトの血清を注射し発生した抗原抗体反応を100とする
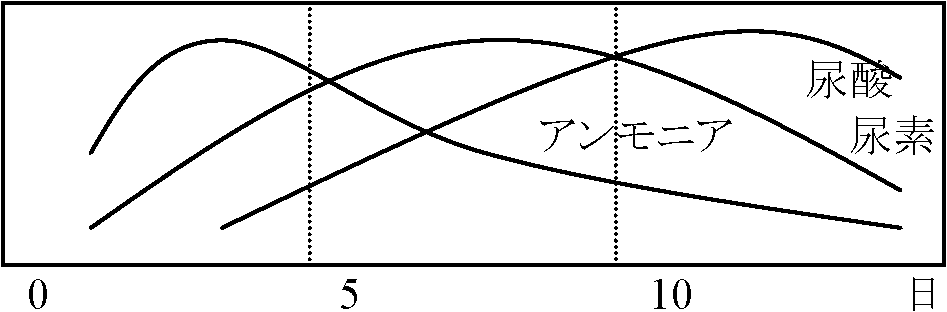
ヒト ゴリラ クモザル ウシ イヌ ウサギ a) 窒素代謝産物排出 NH3: 硬骨魚類 尿素: 水に可溶 → 両生類、軟骨魚類、哺乳類 (哺乳類の場合、進化途中で胎生となり母親の方に排出物を渡せるので尿酸に変える必要がなくなった) 尿酸: 水に不溶 → 爬虫類、鳥類、(昆虫類)。陸生で卵生のものに発達
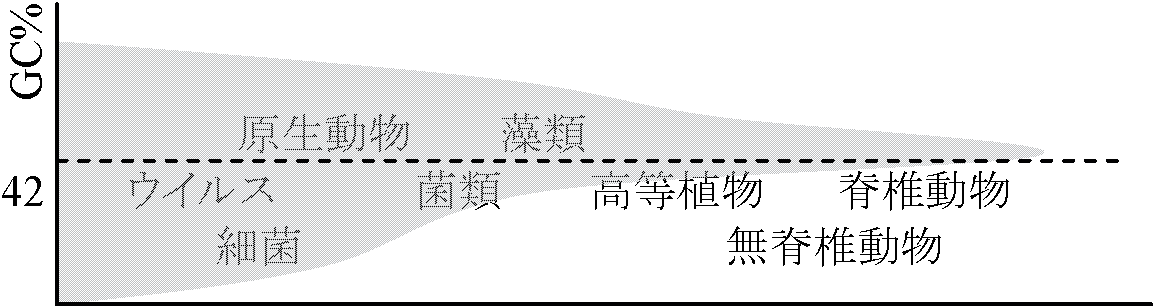
相対変化 G, Cの多い方が密度は高く密度差を利用して測定 → 混ぜ7 M CsCl, 超遠心(40000/min, 20-60 hrs) = density gradient形成
密度違えばDNA解析できる チトクロームc: 104個のアミノ酸からなる1本のポリペプチド鎖。全生物で共通なのは35個のみで、この35個所の立体構造がチトクロームcの機能を保存している。[仮定] 類縁関係近いものほどアミノ酸配列類似性高い → 系統樹(分子系統樹)作成 Ex. チトクロームcはアミノ酸1個の変化に約2000万年(逆算可能) その他の各種タンパク質による系統樹が作成されている 変化: DNA塩基 → アミノ酸 → タンパク質 → 機能・特性 → 生物種 5) 分類学a) 中間型存在 カモノハシ: 爬虫類-哺乳類の中間(卵生、総排出口もつ単孔類) シダ種子植物 Ex. ソテツシダ シダ – イチョウ・ソテツ(動く精子) – 裸子植物 – 被子植物 シーラカンス: 魚類-両生類の中間b) 遺存種 relic: 生きた化石 – 環境変化に耐え、遅い進化速度により過去の地質時と殆ど同じ体制で現存 化石の代わりに生体を詳しく調べられる
Ex. 動物: カブトガニ、ムカシトンボ、シーラカンス a) オーストラリア大陸 新生代第三紀始め頃アジア大陸から分離。当時の哺乳類は単孔類、有袋類。他大陸では有胎盤類が表れ単孔類、有袋類が絶滅したが、オーストラリア大陸では有袋類が適応放散した b) ガラパゴス諸島: 固有種 Ex. フィンチ、ゾウガメ、ウミトカゲ(イグアナ) 適応放散の生態的条件 A) 十分に大きな面積 + 地形的多様性topographic relief → 隔離形成 B) 分散dispersionの困難性 赤の女王仮説 Red Queen hypothesis
科の淘汰確率はランダム 被食回避 predator avoidance直接防衛: 自身による防衛(s.s.) + 耐性・補償作用(s.l.)化学的防御
毒: アルカロイド・シアン配糖体・カラシ油配糖体 生物的防御 間接防衛: 多種を利用した防衛 Ex. 内生菌、捕食者(寄生者)、昆虫(アリ) Ex. [植食性昆虫] ↓(摂食) ↖(麦角アルカロイド) [禾本 ------------ 麦角菌] |
生存競争 struggle for existenceダーウィン進化説の中心的概念 (☛ 競争)同種・異種個体間で限られた資源を奪いあう競争 - 勝者の遺伝子が増える 自然状態での競争Pianka: 競争重視 → 存在の証拠を得る困難性= 現在の自然環境ではある程度競争が済んでしまった
適応の証明方法直接法(再現法)現存遺伝的変異: 種内2変異型比較 → 高成功率の方を知る Ex. 工業暗化
表現型の違いは遺伝型によることが前提
1967 Tinbergen: ユリカモメ親が孵化直後に卵殻を捨てる行為
卵はあるが卵殻が除去されない人工巣作る → 比較法(ある意味常道): 異なる選択圧に晒されている種(個体群)の比較
≈ 相関論 ≠ 因果論
Ex. 食虫性コウモリのエコロケーション echolocation
出す音波の波長は虫より小さくないと反響しない = 高音必要
選択圧強 - 設計特性明瞭 → 高度音響設計できなかったコウモリ絶滅? 適応度(繁殖成功度) fitness古典的適応度, WA = (個体の)適応度
= その個体Aが次世代に残す子供数(の期待値) 包括適応度 inclusive fitness, Wl遺伝子コピーは親子以外の血縁個体同士も共有することを考慮= WA + Σi=1n[(近親者iの適応度, Wi) × (A近親者の血縁度ri)], i ≠ A Def. 利他行動(現象) altrism: 個体が自己適応度下げ他個体適応度上げる利他的形質判断法 Ex. その形質が自己防御に役立っているかを知る
r, ri: 個体間血縁度 relatedness
同世代に属する個体間の利他的相互作用のみに適用
母系社会: 雄は一時的に群れに滞在(雌占有)
乗っ取る群れの雄と闘争 ⇒ 雄同士協力(連合) = 乗っ取り成功度↑ 2頭 = 23/39, 3頭 = 11/12, ≥ 4頭 = 9/9
+ 占有期間は連合が大きいほど長くなる 適応度: 単独雄 × 4 = 連合雄 → 進化可能 Case. Aとその妹2個体: Aの利他行動 → WAがcだけ減り、Wiがbだけ増える
この行動の進化条件 ⇒ br – c > 0 利他行動が進化するさい満足する不等式
同父母兄弟姉妹間: b/c > 2
母(女王ハチ)♀ |A|B|─┬─ 父(雄ハチ) ♂ |C|
_______________│ │__│交尾
息子 ♂ |A|B| |AC| |BC| 娘 ♀
娘間血縁度: 父方遺伝子は必ず持つ/母方遺伝子は確率1/2で共有 → 1/2 + 1/4 = 3/4 母娘間血縁度 → 1/2 ⇒ 娘にとって、母より姉妹の方が血縁度高 → 姉妹を世話すること進化可能 Cp. 親の操作説(Alexander 1974, 1991): 母親が子の性・不妊雌を決定 – 社会性進化ではハミルトン説より有力 Case. ヘルパー helper: 兄弟姉妹の世話 + 自身の繁殖活動(一時的に)放棄兄弟姉妹の適応度高まれば進化しうる(現在否定的) Case. 子殺し(同種内殺戮): 自身の子を殺す場合と他人の子を殺す場合
Ex. アカオザル、カオムラサキラングール、ゴリラ、ホエザル、ライオン → 全て実質的には一夫多妻
メスが子殺しを許容する対抗進化が起こらなかった理由は説明不可
餌資源乏しい → 弟妹を巣から排除するか殺してしまう = 群の2個体以上のメンバーを1単位に淘汰 → 淘汰が群レベルで働く
デームdeme: デーム間淘汰 interdemic selection 最適戦略 (最適化理論)Def. 最適性 (仮定: 長期にわたり環境は安定/一定 - 2つの意味 ⇒)1) 長期的: 一生の間に最も多くの成体に達した子を残す 2) 短期的: 一定時間内での最適行動 Ex. ネズミ採餌行動
全く別物 → 混同してはならない (一緒にするのは誤用)
適応戦略に応用: (結果として)効率良いものに全力尽くす ハミルトンの法則と包括適応度= 利他行動進化の条件r > c/b ⇒ 量的遺伝形質の集団平均値G(集団中の利他者割合)増加条件 r: 血縁度(近縁係数) relatedness, c: 損失cost, b: 利益 benefit R(G) = W0 – cG + bG·r = W0 + (br – c)G
G: 形質値 = 利他行動の発現確率 a) br – c > 0 → r > c/b ⇒ ハミルトンの法則: G大きい個体ほど高い包括適応度 → G増大 b) br – c < 0 → r < c/b ⇒ G小さい個体(= 利他的傾向弱い個体)ほど高い包括適応度 両賭戦略(二股戦略) bet hedging子の再生産成功率が予測不可能(Ex. 撹乱地、荒地、砂漠) → 不確実な状況ではリスク分散が有利⇒ Def. 両賭戦略 Ex. カダヤシ: 水位変動 小 → 大
体小/繁殖投資大 = 親生存不安定 |
ハミルトンの拡張法則: 行動進化条件式α + βr > 0, r > -α/β or r < -α/β → α + βrは正なら利益、負なら損失
α: 自らの適応度変化 α > 0, βr < 0 利己行動: 血縁間個体間では不利 α > 0, βr > 0 両得行動: 一見利他行動に見えるるものにこの可能性 α < 0, βr < 0 両損行動: 現実的に進化し得ない Ex 1. 最適採餌戦略(最適採餌理論) φ = (1餌平均カロリー量)/
[(1餌処理平均時間) + 餌処理時間: 餌に出会ってからそれを捕らえ食い終わるまでの時間 Ex 2. 卵サイズφ = R/ρ·s(ρ)
φ: 次世代の親になれる卵数 最適戦略は dφ/dρ = R/ρ(-s(ρ)/ρ + s'(ρ)) = 0 で求められる 自家中毒 (自己中毒 self-intoxication)種内要因(有害物生産等)により増殖率が(負に)影響される場合 Ex. 排泄物仮定: 有害物生産量は個体数に比例 A: 効果が一定で持続する場合時刻tでの増殖率: 蓄積された有害物質からの影響を考慮
(τ (τ < t) → τ + dτ)間の生産有害物質がtにおける増殖率への効果 ⇒ f(t - τ): (毒性)残存効果 after-effect dN/dt = [ε - λN - ∫0tf(t – τ)N(τ)dτ]NCase. λ = 0, f(t) = c (一定) ⇒ X(t) := ∫0tN(τ)dτ d2X/dt2 = (ε - cX)·(dX/dt) = d/dt·(εX - 1/2·cX2)
dX/dt = -1/2(cX2 - 2εX - 2N0)
α, β: cx2 - 2εx - 2N0 = 0の根
M(t) = ∫0tf(t - τ)N(τ)dτ = ce-νt∫0tN(τ)eνtdτ ⇒ 以下の連立微分方程式ができる
dN/dt = (ε - λN - M)N
(N, M) = (0, 0) 進化的安定戦略 evolutionary stable strategy☛ ゲーム理論(経済学) ゲーム理論 game theoryEx 1. 採餌場所選択ゲーム(Maynard Smith 1982, 巌佐 1990)x1, x2: 2つの採餌場所(i = 1, 2)にいる個体数 (全個体数 N = x1 + x2) 仮定: fi(xi) = ai – bixi (負の一次相関)
xiで個体当り摂食速度は採餌個体数増加につれ減少 Case. N↑: 各個体が摂食速度高い方を選択 ⇒ f1(x1) = f2(x2) (平衡状態)
≡ 理想自由分布 ideal free distribution Case. 協力ゲーム (個体が協力しN個体全体での資源利用効率高める) φ (利用した資源量) = x1f1(x1) + x2f2(x2) N = x1 + x2の下で φ*を求める Ex 2. "どちらの親が子の世話をするか"ゲーム = 最単純ゲーム仮定: 雄雌2個体 → 生まれた子の世話は自分の繁殖成功高まるなら行なう
子の生存率: 両親から世話 = S2、片親 = S1、世話なし = S0
________________メスが子の世話を
両親(進化的安定条件 evolutionarily stable) ⇒ 2つの上式を満たす V: 闘争に勝った時の利得. C: 負けた方の損失
_____________相手が 仮定: コストは時間に比例(ct, t: 待つ時間) p(t) = exp(-c/V)t)c/V 分散と遺伝子フロー dispersal and gene flowEx. Salisbury (1961)
Verbascum thapus: 12 feetで急激に花粉散布量が減少する 任意交配集団panmictic units (≈ subpopulation)と近隣関係 neighborhood 理論的近隣サイズ neighborhood size, Ne = σ (散布距離のSD, standard deviation) × 2 Ex. Phlox pilosa: N (number of individuals) = 75-182, Liatris aspera = 30-191, L. cylindracea = 165, Lithospremum caroliniense = 4 遺伝子フローと連携群(レース)分化 gene flow and racial differentiation1963 Wright: 環境圧を考慮しない場合
N______10____________100___________> 1000 遺伝子フローと淘汰 interactions between gene flow and selectionpanmictic units 小 / envrionmental pressure大 → racial diffferentiation起こりやすい Local race形成は淘汰圧と遺伝子フローの割合による他殖植物でのrace形成 – disruptive selectionが強ければrace形成可能 edaphic factor, biotic interaction (=competition) Ex. Agrostis tenuis (Poaceae) – micro-evolution (Bradshaw 1959)Ex. Potentilla (Rosaceae) – 4つのsubspeciesに分類 死滅分散: 分散 → 生息場所拡大 自然条件では子孫存続可能性のない地域まで世代を繰返しながら分布拡大 Ex. ウスバキトンボ: 高温期に東日本まで世代を繰り返しながら移動 – 冬の低温を乗り切る能力なく結局子孫を残せないまま死滅する |
|
Def. 境界条件 boundary condition: 境界に課される条件 Ex. 条件: 周期的境界、ディリクレ境界(固定端)、ノイマン境界(自由端) 自己組織系へのアプローチ1968 Polanyi Michel: 二重制御理論
= 生物機構は機械と同じであり二重制御の元で働く 拡大二重制御理論 (仮説)複雑構造 (Ex. タンパク質)↑→ 物理化学的制御(物質過程) (Ex. 自由エネルギー最小則 = 熱力学的法則) ↑→ 情報的制御(生命過程) (Ex. アミノ酸配列) 単純構造 (Ex. アミノ酸) → 階層構造: ニワトリが先か卵が先か。個体発生は系統発生を繰り返す |
1971 Eigen Manfred: アイゲンの進化方程式
自己触媒を含む化学反応ネットワークの中から情報の根源である遺伝コードが生まれる → 自己複製系の確立と情報生成は等価 自己複製系誕生 ⇒
"必然" Ex. アイゲン(進化方程式)、伏見(進化分子工学) |
植物界 Plant kingdom1. 核分化a. 原核植物(前核植物): 核-細胞膜区別なく、ミトコンドリア・色素体等が見られない前核細胞からなるb. 真核植物: 核膜がある真核細胞からなる 2. 細胞分化a. 単細胞段階
単独(単生) solitary: 遊泳型 mobile form, 粒状型 cocoid form c. 多細胞段階
単列糸状体 uniseriate filament / 柔組織 parenchymatous tissue 3. 器官分化a. 葉状植物: 器官分化がない = 細菌 – コケb. 茎葉状植物: 根・茎・葉の分化がある。維管束植物そしてまとめることもできる = シダ – 種子 4. 生殖法a. 分裂植物: 分裂で増える = 細菌・藍藻b. 胞子植物(隠花植物): 胞子で増える
b1 造卵器を作らない
生活史型: 減数分裂 (例)
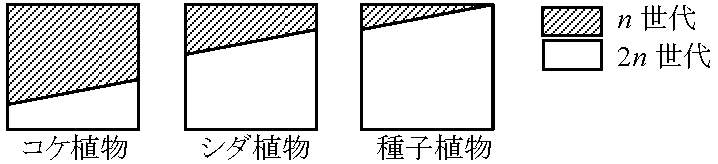
2n > n (コンブ、シダ植物、種子植物)
単相植物 nのみ: 接合子発芽 (アオミドロ、シャジクモ、フラスコモ) |
5. 栄養摂取法a. 独立栄養植物: クロロフィル(同化色素)持つa'. 従属栄養植物: クロロフィル持たない b. 藻植物(藻類): 同化色素を持ち水中生活 b'. 菌植物(菌類): 同化色素を持たず寄生生活 植物系統原始生物: 細菌類様従属栄養単細胞生物 → 細胞壁完成した細菌植物が、一方でバクテリオクロロフィルを持つ光合成細菌が進化。光合成細菌からクロロフィルを持つ単細胞藍藻植物が現れ、核、色素体、ミトコンドリア、小胞体、ゴルジ体等のない前核生物が生じた。前核生物から真核生物が生じたが、まず藍藻類のあるものから、核、鞭毛等の分化により黄色鞭毛藻植物が進化し、一方で無鞭毛の紅藻植物が進化した。細菌植物のあるものからクロロフィルを持たない真菌植物や菌類と藻類の中間型を示す藻菌植物が分化した。他方、鞭毛を持つ細菌から変形菌植物が分化したと考えられる。黄色鞭毛藻植物から一方向に珪藻植物、渦鞭毛藻植物、褐藻植物、輪藻植物が分化した陸上植物の進化系統に関する仮説1) 輪藻植物 → コケ植物 → シダ植物 → 種子植物2) 原始シダ類からコケ植物と種子植物という2系統が分化 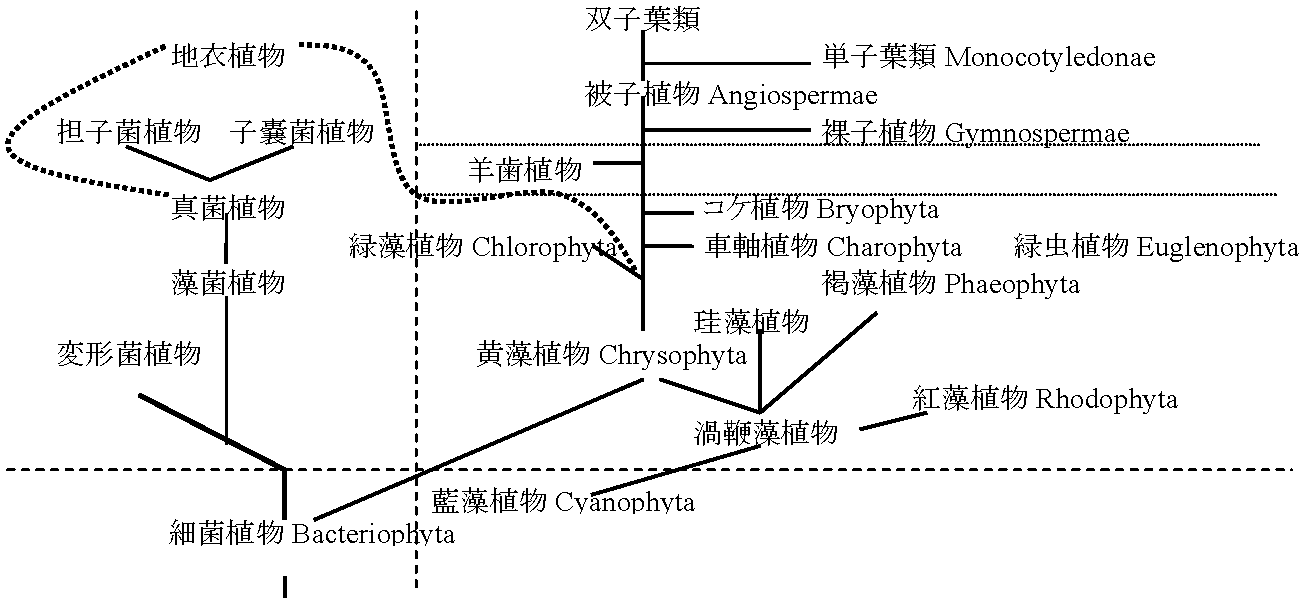
|
| カロチノイド carotinoid | |||||||||||
| 門 | 核 | クロロフィル | 葉緑体 | ラメラ構造 | カロチン carotin | キサントフィル xanthophyll | タンパク 質色素 | 貯蔵物質 | 細胞壁 | ||
| 細菌 | - | - | - | チラコイド | (β) | - | - | - | C, P | ||
| 藍藻 | - | a | - | 一重 | β | - | - | [m] | ++ | 藍藻デンプン, 藍藻粒 | 種々粘液 |
| 炎藻 | + | a, c | + | β | - | - | [d] | + | クリプトアミロン, F | C | |
| 黄藻 | + | a, c or e | + | 三重 | (α), β | (l) | (v) | [(f)] | - | ロイコシン, ラミナリン, F | P(珪酸質) |
| 褐藻 | + | a, c | + | 三重 | α,β | (l) | (v) | [f] | - | マンニット, ラミナリン, F | アルグロース |
| 藻菌 | + | - | + | - | - | - | - | G | C, T | ||
| 古菌 | + | - | - | - | - | - | - | G | C, T | ||
| 粘菌 | + | - | - | (β) | - | - | - | G | - | ||
| 真菌 | + | - | - | (β), γ | - | - | - | マンナン、G, F | T | ||
| 紅藻 | + | a, (d) | + | 一重 | (α), β | l | - | ++ | 紅藻デンプン, F | C | |
| 緑虫 | + | a, b | + | 三重 | (α), (β) | l | - | [e] | - | パラミロン | - |
| 緑藻 | + | a, b | + | 多重 | (α), β | l | v | [(e)] | - | S, (F) | C, P |
| 輪藻 | + | a, b | + | β, γ | l | v | - | S, F | C, P | ||
| 蘚苔 | + | a, b | + | グラナ | α, β, γ | l | v | - | S, F | C, P | |
| 羊歯 | + | a, b | + | グラナ | α, β, γ | l | v | - | S, F | C, P | |
| 種子 | + | a, b | + | グラナ | α, β, γ | l | v | - | S, F | C, P | |
(): 微量。[ ]: 特殊。l: lutein, v: violaxanthin, d: dinoxanthin, f: fucoxanthin, e: euglenarhodon. G: グリコーゲン, S: デンプン, F: 脂油, C: セルロース, T: キチン, P: ペクチン
藍藻: 藍藻素 phycocyanin を含む-菌類
小史田中芳男(1838—1916): 文部省博物局1875 田中芳男(訳纂)「動物学初篇哺乳類」 1877 東京大学創立: 理学部動物学教室
初代教授 Morse, Edward Sylvester (1838-1925) 分類原始動物 → 鞭毛虫類様原生動物 → 原生動物/有鞭毛室海綿動物嚢胚期以上の発生 = 二胚葉性器官分化した腔腸動物 (間に中生動物) 二胚葉性腔腸動物 → 三胚葉性動物(2系統)
発生単細胞 ___ 原生動物 protozoan多細胞 2胚葉 ___ (海綿動物)、腔腸動物 3胚葉 旧口動物 原体腔 ___ 扁形動物、紐形動物、袋形動物 真体腔 新口動物 脊索できない ___ 棘皮動物 脊索できる 羊膜なし ___ 魚類 (fishes)、両生類 (amphibian) 羊膜あり 胎盤なし ___ 爬虫類 (reptile)、鳥類 (birds) 胎盤あり ___ 哺乳類 (mammals) → 人類 (human) 細胞分化原生動物 ___ 単細胞後生動物
中生動物 ___ 組織分化がない 発生段階受精卵段階 ___ 原生動物桑実胚段階 ___ 中生動物 嚢胚段階
二胚葉(内外胚葉)段階 ___ 側生動物(海綿) 中胚葉の出来方
原中胚葉細胞幹: 原中胚葉細胞が発達し中胚葉になる |
原口と口の関係
旧口動物 (前口動物 Protostomia): 原中胚葉細胞幹 → 原中胚葉細胞(端細胞)が細胞分裂により中胚葉を形成する。原口 → 将来口(前側) 例外: 触手動物は前口動物で原腸体腔幹 体腔: 間充織 = 内外両胚葉の間にできる結合組織。未分化星状細胞が疎らに集まってできている
原体腔: 卵割腔がそのまま残ったもの。間充織をためた狭い体腔をもつ – 扁形・袋形(紐形・輪形)
脊索できない ___ 原生-節足、棘皮 (無脊椎動物)
脊索は一生存在 ___ 原索動物 (無脊椎・脊椎動物)
無腸動物: 消化管もたない = 細胞内消化 Phylum Vertebrata 脊椎動物門 vertebrates (chordates)内部体節制internal segmentation: 骨格・神経・筋肉 - 骨格vertebra (内骨格 endoskelton)を持つ頭部集合cephalization: 骨格・神経は頭部に集合 脊椎動物 Subdivision Vertebrata無顎類 Agnatha (過去魚類に含む): 顎骨がない円口類 Cyclostomata or Cyclostomi
ヤツメウナギ目 Petromyzontiformes
板皮類 Subclass Placoderm: 顎骨もつ原始的脊椎動物
両生類 Subclass Amphibia 恒温動物 homeotherms vs 変温動物 poikilotherms恒温動物: 外界温度変化に関わらずほぼ一定の体温を維持する動物
体温低下時期(Ex. 冬眠)がある
温度が安定な環境では体温変化は小さい 外温生物 exotherm vs 内温生物 endotherm外温生物: 体温調節の熱を外部熱源に依存する生物内温生物: 体内で熱を発生し体温調節する生物 |
|
生物体の構造と機能を発生過程を含め明らかにする → 生物体の形 - その意味(機能、発生、適応、進化)を明らかとする学問 肉眼解剖学 gross anatomy: 実体顕微鏡程度を補助に用いたメスやピンセット等の道具を使用する生物解剖 顕微解剖学 microscopic anatomy: 組織包埋しミクロトーム等器械を用い切片を作り顕微鏡観察 系統解剖学 systematic anatomy: 生物を骨格系・筋系・神経系・脈管系等の系統別に分け、各系統を記述 |
人体解剖学 human anatomy: 特にヒトを対象 局所解剖学 topographic anatomy (応用- applied -, 外科- surgical -): 生物の特定領域に全系統を記述 発生学 embryology: 発生の時間経過に伴う個体形態の成長・変化を扱う |