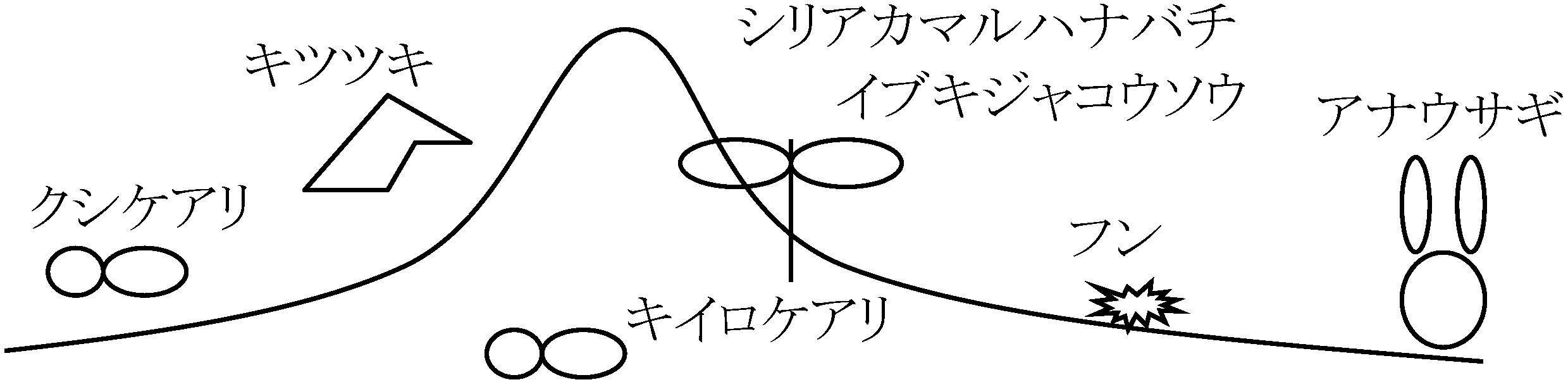(2025年6月16日更新) [ 日本語 | English ]
HOME > 講義・実習・演習一覧 / 研究概要 > 小辞典 > 生物学 > 生態学
生態学とは What is ecology?生物と環境又は生物と他の生物との関係を研究する学問 (Haekel 1866)生態系の原理と概念個生態学 autoecology: 生物個体あるいは単一種の研究
種生態学 genecology
生態系生態学 ecosystem ecology [日本の植物学者] 日本生態学史 (history of ecology in Japan)1885-1928: "Ecology of plant" (Warming 1895)日本に紹介三好が用語「生態学」を提唱(原語がoecologieかbiologieかは不明) 1945-1954: 鈴木時男・細川竜平・正宗玄: 外地から引上げ = 広く植生観察
→ 日本の植生は立体的に把握 |
☛ 講義: 生態学, 環境保全学特論, 生態系論 A / B |
|
材料区分: 動物生態学 animal ecology, 植物生態学 plant ecology → 生態系 ecosystem = 植物 plants + 動物 animals (+ 微生物など) であり便宜的 __________| 自然誌学 Darwin__________| 関係生理学 Haeckel __________| 適応生態学 Stohl 進化生態学 -| 生態地理学 Raunkiaer - 植物生態学 __個生態学 -| 生物生態学 Clements - 生物地理学 - 群落構造論 環境生態学 -| 生態系生態学 Odum - 遷移論 実験生態学 -| 応用生態学 Ellenberg |
応用生態学農業生態学: 農作物品種改良・栽培管理、農耕地関連土地利用等の諸技術を生態学的手法で研究→ 農業生態系 agro-ecosystem: 農作物栽培により形成された生態系 生態モデル ecological model言語モデル word model≈ 記述モデル descriptive model: モデル化に必要な準備段階数学モデル(数理モデル) mathematical model決定論的モデル deterministic model; 予測値は点確率論的モデル stochastic model: 予測値は確率分布 |
|
Darwinian理論を動物社会に導入 → 動物生態学始まる Elton, Charles Sutherland 1900.3.29-1991.5.1 (England)
全く異なる群集に類似点存在 → 系統進化・分類上異なる種でも同じ地位を占めることはある 目的: 動物行動の機構を理解 行動の起こる仕組: 鍵刺激、行動解発機構、発現方向制御、行動プログラム Ex. トゲウオは「赤色」を成熟雄と認識し縄張から排除を試みる 行動 = 生存価 + 個体発生 + 進化(種に固有な行動 → 遺伝的/生得的) Def. 習性 habit: 個体群中に定着した行動様式≠ 癖 quirk: 個体に限られたもの Def. 種特異的行動 species-specific behavior: 種が遺伝的に持つ特異行動 |
動物生態学方法論= 個体群生態学population ecology (autoecology) ☛ 個体群密度測定法
数 = 個体群サイズ: ある種の生物がある地域のある時点で生活している"数"(個体数)を調べ、その変化要因を明らかにする(伊藤 1983)
Ex. アナウサギ・ネズミ → キツネ, カサガイ面積(岩固着力) → 水鳥の嘴, 血球サイズ(7-18 μm) → ツエツエバエ 生得的(1) ≈ 遺伝的
発達的に固定された行動 Ex.
本能的行動パターン Instinkthandlungen |
1. コミュニケーション communication視覚信号(遠距離や障害物ある所に不向き) + 聴覚信号 + 臭覚信号進化法則a) 信号祖先型存在b) 信号祖先型は伝達信号であるとは限らない c) 自然淘汰が働けば信号はより効果的なものに進化 d) 祖先型を同じくする近縁種は類似行動が見られる Def. 儀式化: 信号機能ない動作や別意味な動作が独立した信号へ変化過程 Ex. おどし、服従、社会的接触、求愛 2. 群れ vs 縄張群れ: 他個体と行動をともにする ⇔ 縄張: 他個体を排除する同一個体群内空間利用生息地 habitatはpatch状かclump状に分布移動力 → 縄張
移動の程度 – 時期・距離・場所等の特性 Ex. 鳥の渡り、魚の回遊 行動圏 home ranges.l.: 個体群か群れの空間のカテゴリー (total range: 一生涯に動く全範囲)s.s.: 一個体が主に動く、パトロールするような範囲 (core area: 塒の周辺)
食物や配偶者を求める活動範囲 = 他個体侵入 → 目立つ防御行動なし 逃走距離(逃避距離) flight distance: 捕食者接近時に逃げ出す最小距離 社会距離(社会的距離) social distance: 個体群間の保ちうる最小の距離1974 Seghers (Wilson 1975中引用)
西インド諸島トリニダッド島におけるグッピー Poecillia reticulataの行動 集団産地 Guayamare Aripo Petite Aripo Paria (滝の下) Curucaye (滝の上) 主捕食者 大形魚類 大形魚類 メダカ(多) メダカ(中) メダカ(少) 群泳行動 強度に発達 強度 中度 弱度 なし → 捕食者に対する個体防衛 → 遺伝的形質(継代飼育で証明) Def. 縄張制 territoriality: 個体群構成個体が同種他個体に対し一定の生活空間を占有し侵入者を排除しようとする働き = 空間独占性 / 安住性Def. 縄張 territory: 縄張制がある時、その個体の勢力圏
先住効果 prior residence effect: 先住者の方が縄張闘争に有利
Ex. アユ: 行動圏 ≈ 2-3 m2位 > 縄張 ≈ 1 m2 (個体サイズ関係) 帰家行動 homing: 特定の場所の記憶必要 - 中枢神経発達 縄張形態: 個体、ペア、群れ 仲間識別できる高等動物、特に脊椎動物に見られるが、アメンボやシオマネキ等の無脊椎動物にもあるEx. arthropoda, vertebrate, メダカ・トゲウオ・アユ等の魚類, アリ・スズメバチ等の社会生活をする昆虫
Ex. アユ: 川底の石に付着する藻類採餌 (川遡上する初夏には数匹から成る群行動) (Wilson 1975, 伊藤 1978) 縄張の分類
諸説あり (Allee et al. 1949, Hinde 1956) 縄張の機能目的: アユ縄張 = 食物確保 vs トンボ雄縄張 = 主に配偶者としての雌確保
Ex. ハッチョウトンボ雄: 湖沼の周辺や湿地に縄張を作り、他の雄が侵入すると追い払う。しかし、雌が入ると交尾し、その雌が産卵を終えるまで警戒飛翔を続ける。雌が他の雄と再び交尾すると、最初の雄の精子が卵の受精に使われなくなるからである i) 個体維持に関する機能
個体群を空間的にうまく配置し、個体群密度を保ち個体間競争を緩和 - 密度依存的作用調節
つがい形成と維持を容易にする - 交尾失敗時に再婚できる 2) 二次的機能
個体群一様分散 → 好適な住み場所調節し新しい住み場所へ植民促進 縄張成立に必要な餌量を予測: E + T < aP + e(1 - a)P
E: 生活に必要とする基本損失
P: 採餌範囲における餌の全生産量 e(1 - a)P: 縄張を作ることにより得られる利益
(1 - a)P: 縄張により余分に得られる生産量
Case. 防衛完璧 → e = 1 (全侵入者を排除) ⇒ E + T < P 実測値と概ね一致 ∴ 縄張が餌確保(採餌)に有効 鳥の縄張: ミソサザイの一種(Howard 1907-), スズメの一種(Nice 1931)縄張は細区分され繁殖期にだけ見られ、侵入者を攻撃するのは主に雄 縄張は個々の動物の生活環境で目立つもの Ex. アユ: 水底の石や藻が目印 図. 縄張性の進化に影響をおよぼす諸要因 (Brown 1964) 移動 migration個体群はいつも同じ生息地に留まると限らず、鳥・魚では、群れで長距離移動することもある |
☛ 渡り(鳥類の移動)回遊 recurrent migration魚類等の水生動物: 食物、適温、産卵地を求め移動すること(s.l.)1) 適当な温度や食物を求めて移動 Ex. イワシ、サバ、ニシン、ブリ 2) 回遊環 loop (回遊 s.s.): 産卵地と生息地異なる 海洋回遊: 回遊が海だけで行われる 河川回遊: 回遊が淡水だけで行われる 通し回遊: 川と海の間を行き来 → 回遊魚: 通し回遊をする魚類 遡河回遊: 川を産卵場、海を成育場として利用 Ex. サケ科魚類 Ex. サクラマス: 降海型、降湖型、河川残留型、河川型、湖沼型、陸封型がある
降河回遊: 海を産卵場、川を成育場として利用 Ex. ウナギ、カマキリ サケ母川回帰 → 幼魚時代の川の匂いを数年間に渡り記憶している 河川陸封型(河川残留型): 本来は河川と海を回遊(一往復)していたが河川で一生を過ごすようになったものEx. サツキマス陸封型 = アマゴ(矮小化)。サケ科パーマーク存在 湖沼型(降湖型): 河川と湖沼で特異な形態が見られる群生活の発達 (development of group life)Def. 群れ: 個体の密な集団 → 内部構造は種により様々Ex. アリのコロニー colony: 高度分業制を示す母子・姉妹集団 群れswarm: (動いている)昆虫の群れ、(分巣時の)ミツバチの群れ、(混乱した人や動物の)群れ Ex. イワシ・サンマ群れ school: 血縁関係ない多個体が作る単なる移動集団
個体は、集団を作りより効果的に獲物獲得でき、逆に天敵から逃れる 群れの1羽がオオタカ接近に気づけば、全個体が逃げられる + 配偶者の獲得や子育てが容易になる 群れの機能 = 組織化 organization による機能
組織化形態で群れの性質異なる Ex. つつき順位、リーダー制
A. 順位制(オーダー) dominance hierarchyDef. 順位 (dominance) rank: 個体群構成個体間に強弱 → 生活上で序列Def. 順位制: 順位関係が個体群内で優位者のみに有利ではなく全体秩序維持に役立つ制度となっているとき → 機能: 群内個体間関係が安定し無用の争いが減ることが最大 → 攻撃性 aggressiveness と関係 = エネルギーの不必要な消耗や傷つけあいによる群れの劣勢化を防ぐ Def. 優位: 群れ内部で順位成立 → 勝者の地位 ⇔ 劣位1. 絶対順位 peck-right: つつき返しがない 1913/1922 Schjelderup-Ebbe: ニワトリ集団 - つつきの順位 pecking order
絶対順位: pecking right - 直線的 linear Ex. A → B → C → D …
1羽は全てに優位、1羽は一方に劣位を一方に優位、1羽は他2羽につつかれてもつつくことはない
相対順位: pecking dominance - 3すくみ Ex. A ↔»» B ↔» C …
新しいニワトリを加えると、互いに一通りの闘いをやり順位を決める A B C D E Ex. 順位制: 魚類、鳥類(ハト)、サル・シカ・ウマ等ほ乳類、アユ(魚類)、アシナガバチやアリ(社会性昆虫) 1922 坂口総一郎: Macropodus opercularis(トウギョ)の巣防衛行動 戦闘は♂: 多くの該魚を」飼育 → 同一水槽由来の個体は戦闘なし ∵ 既に順位が形成されていた 2. 相対順位 peck-dominance: 互いにつつきあう Ex. モリバトDef. ディスプレイ(誇示) display: 他個体に対し視覚・聴覚・嗅覚等を通し、自身の状態・意図・優位性等を誇張し伝える動様式
儀式化、種固有性、リリーサー機能 1954 河端: Aplocheilus latipes (メダカ)に順位
一方的攻撃で反撃は殆どない - トップとそれ以外の闘争が主 B. リーダー制 leadership organization群れ全体行動を指揮するリーダーleaderによって組織化されること外敵防衛と食物探索の点から経験豊富な老個体が、また子保育する関係から雌個体が選ばれる傾向 Ex. 脊椎動物によく見られる。アリ等の集団も移動時にリーダーが存在するといわれる 動物によってリーダーは順位最優位者と限らない。ニワトリでは中位の何羽かが集団でリーダーの働きを示す。ヒツジやヤギでは雌親がリーダーになることが多い。リーダーは群行動でよく見られ、シカ類では敵を始めに発見した個体が尾を立て走ると、それをリーダーに全個体が一斉に走り去る
ヒトリザル 強力な数個体がボス → 最優位者(ボス中のボス)が君臨すること多 群全個体に順位があり、2個体の中間に餌を投げると高順位者が食べる。高順位者が十分飽食すると低順位者上に乗り優位者を誇示した(マウンティング, 背乗り mounting)後、劣位者は食せる。マウンティングの他に、プレゼンティング(尻をつき出す交尾姿勢)、グルーミング(毛繕い)等の行動で、個体間優劣関係を確認し合う安定した順位制が成立している ボス周辺に雄雌と乳飲み子が位置し、子は育つと若雌は次第に雌親群へ近づき、若雄は周辺部へ出て行く。周辺に位置するのが普通雄で、その中で力ある者がサブリーダー(ボス見習)となり、その内側に位置しそれぞれ縄張を作る。ボスは雌や子を保護すると共に、雌間の喧嘩仲裁や、雌や子の食物をとった若雄を群から追放したりする。群全体の行動はリーダーが支配する
┌ 個体の分散 → 縄張(テリトリー) 群れの調節 regulation of group structure交尾行動 (≠ 妊娠・出産) + (餌)資源量 → 群れ調節に関係 ☛ 個体群増殖最適群れサイズ optimal group size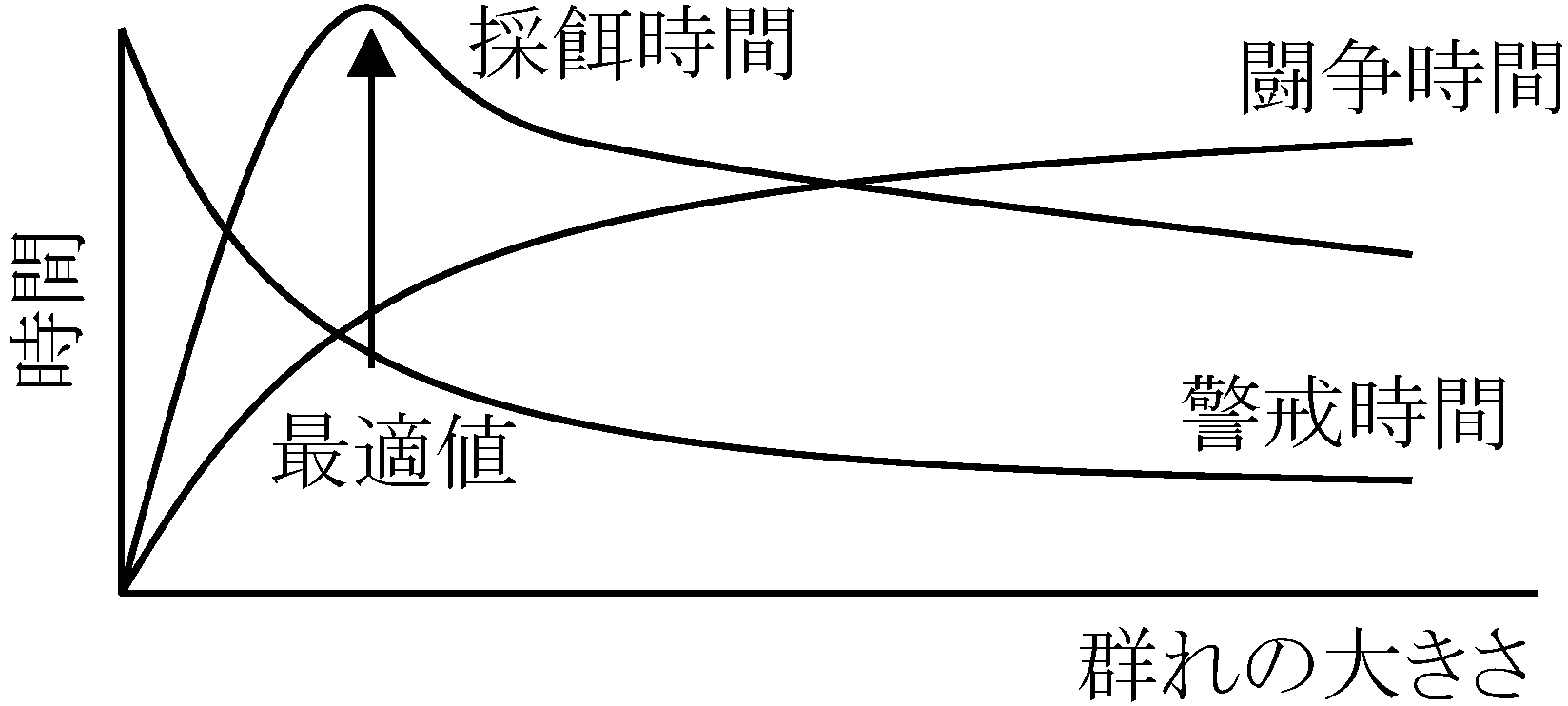 ☛ (伝播 + 個体群周期)
☛ (伝播 + 個体群周期) |
|
Def. 個体群内で全体生活や生殖に影響もつ個体間相互関係に基づく秩序
社会形成 → 哺乳類や爬虫類・魚類の移動や渡り、回遊時に見る Ex. 高等哺乳類: リーダー持つ社会形成 Ex. オットセイやアシカ: 強力な雄を中心に縄張中にハレム形成
他雄を追い出し雌と交尾し子を生ませ育てるのを助ける
ニホンザル社会: 群の縄張は明瞭 Ex. ニホンザル: 順位制、縄張制、リーダー制を基盤に社会形成 生得的社会性: 社会性昆虫 social insect - 雌親(女王)中心に個体間分業 ミツバチでは女王となるべき個体が同時に表れると双方が対決し一方が死ぬまで闘い勝者が社会形成の中心になる。一定期間中に女王候補個体が表れないとハタラキバチ幼虫にロイヤルゼリーを与え女王を作る 社会構造発達と繁殖戦略a) 高等な哺乳類に見られる社会生活発達
┌ 類家族社会 - ゴリラ ┐ カースト: 分業固定的 - 状態変化しても各個体の役割変わらない
繁殖カースト – 最も発達したものでは労働能力損失
女王バチ queen 1個体 ♀ 膜翅目(アリ・ミツバチ・スズメバチ)、シロアリ目、半翅目(アブラムシ)
ミツバチダンス – 情報伝達
社会性昆虫の行動が単独ではなく集団の存在に誘発・調整される
少個体数で発現 (↔ マス効果 mass effect: 多個体、効果負)
Def. ステイグマジー stigmergy: 環境に残る痕跡(Ex. フェロモン、構造物)が他個体の行動を誘導 亜社会性真社会性に至る手前の段階と考えられる原始的亜社会性: 餌を置いた産卵巣穴を閉じる(一括給食) → 幼虫の間給餌を行なう(随時給食) 進歩的亜社会性: 子供が成虫になるまで母親が生存し、子育てをする Ex. ヒメツヤハナバチモドキ: 母娘二世代の成虫共存 = 娘が母に協力
→ 娘が卵を産まず労働だけ行うハタラキバチにカスト分化 = 真社会性
ホイラー説
ミッチナー説 (ホイラー説否定していない) 父親のみによる保護 vs 母親のみによる保護 vs 両親による保護 ♂: 一匹の子に投資する資源少/自分の子である保証はない →より多くの交尾機会を得るよう進化 ♀: 卵子への投資量は精子よりはるかに大/確実に自分の子 →自分の子を安全に育てるよう進化 ⇒ 雄と雌の闘争conflictの結果 → 子育て様式決定(Trivers 1972)帰結できる動物全体での子育て傾向
→ 一夫多妻多 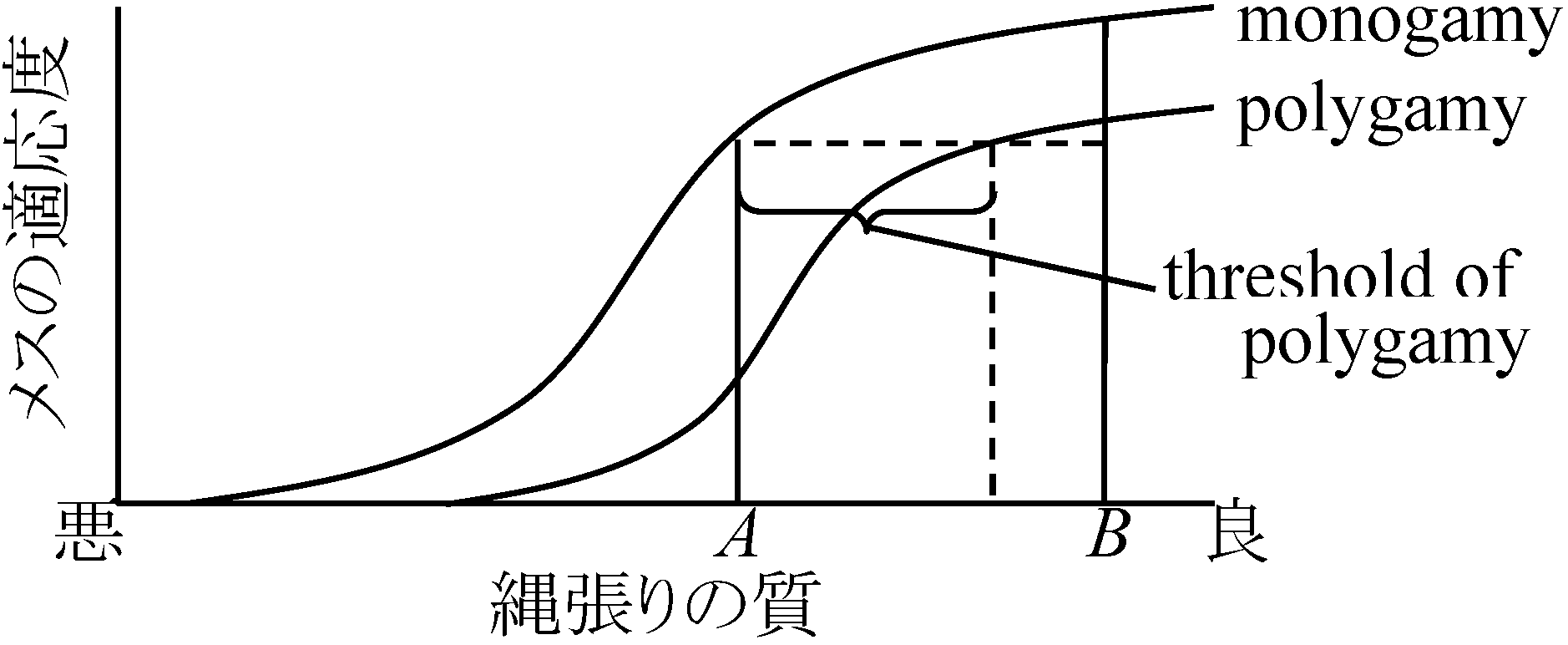 図. 一夫多妻成立を説明するOriansモデル。♂が占める縄張の質が"複婚の閾値"より離れていると♀はAという悪い縄張を占めた♂の第一夫人よりもBというよい縄張を占めた♂の第二夫人のが適応度は高くなる。適応度は、その♀の卵から育った若鳥の数で表わす
図. 一夫多妻成立を説明するOriansモデル。♂が占める縄張の質が"複婚の閾値"より離れていると♀はAという悪い縄張を占めた♂の第一夫人よりもBというよい縄張を占めた♂の第二夫人のが適応度は高くなる。適応度は、その♀の卵から育った若鳥の数で表わす |
Case. カワスズメ科 Cichlidae Ex. Boulengerochromis microlepis (桑村 1988) 沿岸部集合 → ♂♀ペア → 巣(岩場-砂場境, 深3-25 m湖底) - 放卵 → 子育て開始
♀ 水送り(酸素供給), 卵掃除 ↔ ♂ 周囲で捕食者警戒 w/♀
タンガニーカ湖棲息Cichlidae魚類1/3 = 見張型保護(他地域同傾向) ♂ (巣から少し離れ)防衛 ⇔ ♀直接的世話 防衛効果大 - 雄親を除去すると時に絶滅 一夫多妻のカワスズメEx. Lamprologus furcifer: 大型雄が数匹の雌を囲む行動圏 ヘルパーhelper行動確認 Ex. Neolamprologus brichardi: 見張型保護Def. 口内保育 mouthbrooder: 卵や仔魚を口内で保護
科の約1/3の主で行う。内95%の種で♀のみが行う 石斜面に産卵 → 10-20個の卵を雌が咥える(口内保育) → 約2週間後(仔魚 8 mm l)には雄が口内保育(7-10日程度)
口内保育期間中、親も子もは摂食なし(子は卵黄のみに依存) 雌が口内保育 → 雄に交代 + 両親で見張り 托卵 brood parasitism: Synodontis multipunctatus(カッコウナマズ) - 産卵時カワスズメより早く孵化 - 口内カワスズメ孵化個体を餌とし育つ → 口内のナマズは最大で8匹(平均3) - 餌量関係 Case. スズメダイ科 Pomacentridae多: ♂ = 卵見張保護 → 孵化 = 保護終了 ⇒ 仔浮遊期 Ex. (多種) Chromis notata, Pomacentrus coelestis
普段: 餌(藻)のための縄張を♂♀共に形成
♂: 別の♀が来る - 求愛ダンス ⇒ 複数雌の卵を同時に保護 ⇒ 多夫多妻(複婚) - 雄雌の利害対立 Ex. (例外) Acanthochromis polyacanthus浮遊期なし - 仔稚魚を両親が見張り 親: 繁殖期初期に仔攻撃し追出す → 2回目の繁殖成功確率↑ Ex. Dascyllus aruanus: サンゴ(礁) = 隠れ家サンゴサイズ = 収容可能個体数 → サンゴサイズにより行動変化
小 2(匹) = 一夫一妻 ⇒
多くは浮性卵(サイズ ≈ 1 mm、油球により浮く) = 子育てしない ペア産卵 pair spawning vs 群産卵 group spawning ↓ペア産卵(1対1): ♂縄張確保 - 縄張内に1匹の♀が入る
一夫多妻: 縄張には複数の♀が時間を異にして訪れる ⇒ 雌を経験しない生まれつき雄の個体存在 → 小型雄がとる戦略 ↓群産卵: 群れ形成(小型雄個体数は雌の10倍にも) → 1雌が海面に上昇
→ 複数の小型雄が雌を追尾 → 集団産卵と放精 小型雄の繁殖価は雌と大きな違いがない - 共存進化可能 表. 硬骨魚類の繁殖環境と繁殖様式。各タイプの卵を持つ種を含む科数とそのうち子の保護の見られる科数 (桑村 1988)
淡水
海水
沈卵性魚は保護がある程度見られる - 見張り型と体外運搬型が1:3 ⇒ 海の方が卵の拡散性高い - 捕食確率減る 採餌戦略 foraging strategy最適戦略 - 物質・エネルギーで計れる → 利益(benefit, b) – 損失(cost, c)が最大となるとき
利益:成長・個体維持・繁殖
追跡者 pursuer: 容易に餌発見 - 捕獲時間長 Ex. タカ、ワシ、ライオン |
|
= 生態活性物質 ecomone ⇒ 生物間相互作用物質 生産する個体の体外へ放出され、他個体に作用を及ぼすもの
フェロモン pheromone: 同種生物間に作用する物質 フェロモンpheromone1959 Karlson et al. 用語提唱
pherein (Gr. to carry 運ぶ) + horman (Gr. to excite 刺激する) 1. 放出効果 releaser effect Releaser pheromone: フェロモン感受時だけ反応short-term effect。フェロモン受容個体は、直ちに影響(信号)により反応が変わりフェロモン消失により元に戻る。全臭覚器を通じ中枢神経に伝達され行動が起こる a) 性フェロモン sex pheromone (sex attractants): 蛾、マダラチョウ、ハバチ、ヒメカツオブシムシ、キライムシ、ワタミゾウムシ、ワモン、ハイイロゴキブリ等 殆どは雌がフェロモン出し雄反応 (仮説: 雄の方が臭覚器発達する要因)
1956 Baterandt: カイコから雌雄誘起フェロモン単離 |
b) 警報フェロモン alarm pheromone: 社会性昆虫に多 ☛ 防衛戦略
一般に低分子でC = 5-10、分子量 ≈ 100。揮発性多 1971 Wilson, Regnier Jr: 多種多様な警報フェロモン → 総称し警報防衛(alarm-defense system)システムと呼ぶことを提案 c) 道標フェロモンleading pheromone (trail marking pheromone): 社会性昆虫に良く知られるが高等哺乳類にもあると考えられる → これらのフェロモン道標は植物の臭いでもある
Ex. アリ道: アリ尾部末端毒腺dufour glandから分泌 Ex. チャバネゴキブリ: 糞中にrectum pad混入。若齢のものほど集合性高い。シロアリ幼虫でも知られる 2. 引金効果 primer effect → primer pheromone: フェロモン受領後変化が進行long-term effect。社会性昆虫の階級分化に関するものだけが観察されている。内分泌系、臭覚器あるいは味覚器を刺激a) カスト分化フェロモン hierarchy pheromone Ex. プライマーフェロモンprimer pheromone = ミツバチ女王大顎腺から分泌されるqueen substancesが良く知られ同定もされる。この物質は雌に対し働きバチ化促し、雄に対し性フェロモン的役割をする |
|
Def. 都市環境における生物間や生物環境間相互作用を研究
人口増加 → 人間活動-自然環境調和不可 - 問題は形を変えつつ起こる
(中野ら 1974) 変な分野だ 人間と環境原生自然: 環境 Ex. 湿原・森林人為自然: やはり環境 Ex. 水田・農地・住宅地 環境 = 森林 + heath + animals + human 都市環境 urban environment: 都市空間で自然・社会・人工的要素が相互作用し形成される環境
2つのアプローチ 熱汚染: 人間活動により排出される熱 ☛ ヒートアイランド 人間環境としての都市都市は人間にとって万能の地ではなく、成長及び限界の存在を認識都市と共に自然は変化
原初的環境問題: 地下水利用 → 地盤沈下 → 高潮・浸水災害 → 地下水位回復 → 地下水公害発生 都市環境と構造の動態 都市林の機能と効用
防火的効用
都市林の気象及び気候緩和の効用 |
自然林と都市林を比較すると組成的には一般に自然林に比べ都市林は構成種数少ない 緑地(s.s.) 緑(植物)に覆われた部分 Ex. 公園芝生(s.l.) 空間として緑が確保された部分 Ex. 公園全体 - 立体性・連続性・近接性
視覚的緑量: 見た目の緑量(の感じ方) - 緑地面積に直結しない 街路樹、残存樹林、壁面緑化、斜面緑地保全、連続性などが関与 緑地満足度, y = β + a1logx1 + a2logx2 (重回帰式)
x1: 緑被率. x2: 緑視率 人工緑地: 埋立地等、自生樹木のない場所に樹木を導入し形成された緑地 (Wolman 1952) 都市における物質代謝A) 物質移動 (☛ 物質循環)
1) 運搬・輸送 2) 動物輸送 3) 流体を媒体とした移動
1) 物質移動(石油・ガス等) 2) 電線輸送 3) 放射 4) 振動 人間生態学 human ecology人間集団と自然・社会・文化的環境との相互作用 → 生存・生活機構 |