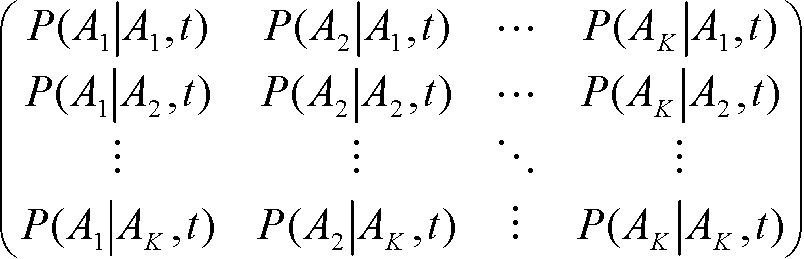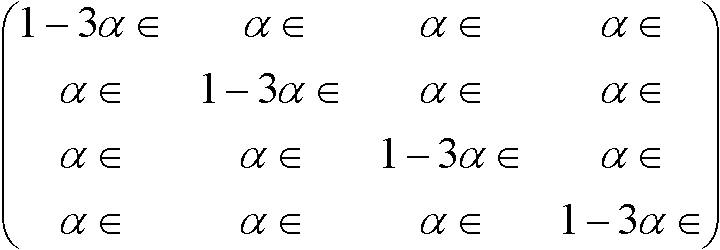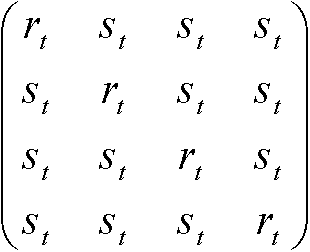(2025年7月14日更新) [ 日本語 | English ]
被子植物分類体系 (taxonomical system)3大体系 (three major syllabuses)新エングラー体系: 構造複雑化説クロンキスト体系: ストロビロイド説 APG体系: 分子系統説 |
[ ベンサム・フッカー方式 ] [被子植物, 裸子植物, 羊歯植物, 化石, 標本整理簿] [植物リスト (plant list)] [図鑑 (taxonomy book), 参考] |
|
1980年代: Cronquist Arthur (1919-1992) ストロビロイド説: 原始的 = 花被・雄蕊・雌蕊多数が軸の回りを螺旋状に配列 = 最も原始的なのはモクレン → 様々に種分化 (キク科が最も進化) |
= 合弁花類・離弁花類の分類群は採用されない ≠ 新エングラー体系
原始的被子植物の形態的特徴を最も良く保存するモクレンの仲間を最初に配列 |
DNAバーコーディング DNA barcoding遺伝子マーカー利用しDNA配列から種を同定系統学 → 系統分岐推定 → 分類体系構築 Ex. ミトコンドリアDNA (特にCOI遺伝子) - 真核生物Ex. rbcL, matK - 陸上植物 形態分類で指摘される問題点(出発点)
重視する特徴により分類結果が異なる + APG (Angiosperm Phylogeny Group)= 被子植物系統作成グループ主導葉緑体DNA解析: 進化研究分野では分類系統の主流となる 形態分類体系と多くの分類群は(当然だが)概ね一致(衝撃的変更もある) APG I (1998): 分類未確定多(暫定)APG II (2003) APG III (2009) APG IV (2016): 64目416科 - 各科の整理順序定める(標本庫対応) APG (2003): 原始的被子植物の共通形質虫媒(甲虫・ハエ類・アザミウマ等が送粉)花弁と萼片の区別不明確 花被片・雄蕊数は3数性 雌性先熟 [ 羊歯植物 ] 羊歯系統分類体系 (PPG)小葉羊歯 + 大葉羊歯2016 PPG I: 基本はAPGに準ずる - 属までの体系
単系統性が第一基準 進化系統樹形態分類学 morphological taxonomy - 外部形態による分類は問題が多い⇒ 形態以外による分類学 Ex. トサボウフウ Angelica yoshinagae (1926) Makino
= A. shikokiana Mak. (イヌトウキ) var. yoshinagae Mak. (1954) Hara 数量分類学 numerical taxonomyEx. Operational taxonomic unit (OTU)を上級概念とする形質評価をしない = 可能な限り主観を持ち込まず分類 (長所かつ短所) 化学分類学 chemotaxonomyEx. 1. 地衣酸: 地衣類の分類に利用Ex. 2. 蓚酸石灰: 単体と集晶の2型
キンミズヒキ Agrimonia pilosa var. japonica – 集晶: 日向 系統樹 (phylogenetic tree)種間進化関係を表わす木: 遺伝配列情報から構成(現在)形態分類と結果が同じとなるとは限らない オーソログとパラログオーソロガスな遺伝子: 種分化により変化してきた遺伝子パラロガスな遺伝子: 遺伝子重複により変化してきた遺伝子 → 進化系統樹を構成する場合は、通常、オーソロガスな遺伝子を利用 ⇒ 階層モデル 系統樹の個数葉の個数と節点の個数の関係(葉の個数 = n)
有根系統樹(根つき木): 根のある進化系統樹 → 2n – 1個の節点、2n – 2本の枝
根は無根系統樹の任意の枝間に設定可能 → #有根系統樹 = (2n – 3) × #無根系統樹
葉 枝(辺): 種の親子関係 枝長: 進化に要した時間 葉: 現在の種に相当する節点 根: 進化の起点にあたる節点 UPGMA法各配列(葉)間距離dijから系統樹構成 – ボトムアップにクラスタを次々とマージアルゴリズム 初期化: 各配列iのみから成るクラスターCiを作る 各配列ごとに葉を作り、それを高さ0の位置におく 繰り返し: dijが最小となるクラスタのペアi, jを求める Ck = Ci ∪ Cjにより、新しいクラスタkを作り、全lに対し距離dklを定義 節点i, jを子に持つ節点kを作り、高さdij/2の位置に配置 kをクラスター群に追加し、i, jをクラスター群から削除 終了処理: クラスタがi, jの2個だけとなったら、根を配置し終了 // 進化の確率モデル分子時計(進化速度の一定性の仮定): 枝長 = 分子時計により刻まれた時間
「任意の葉までの枝長の和は同じ」が任意の節点について成立 → 分子時計の仮定成立 ⇒ ウルトラメトリック 進化の確率モデルP(b|a, t): 長さ(時間)tの枝を辿る事で残基aが残基bに置換される確率 P(x|y, t): 長さ(時間)tの枝を辿る事で配列yが配列xに置換される確率 P(x|y, t) = ΠuP(xu|yu, t) 置換行列(置換確率行列) (K: 残基の種類),
S(t) =
乗法性: S(t)·S(s) = S(t + s) ⇒ |
置換速度行列, R = 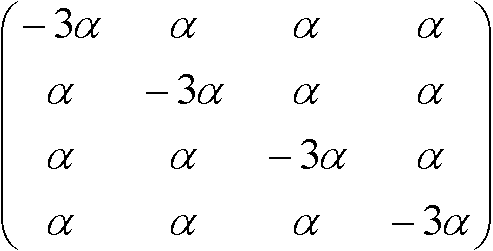
S(t)とRの関係:
S'(t) = S(t)Rよりr* = –3αr + 3αs, s* = –αs + αr Kimura行列トランジッション: プリン(A, G)間、もしくは、ピリミジン(C, T)間の遷移トランスバージョン: それ以外の遷移(トランジションより起りにくい) 置換速度行列: 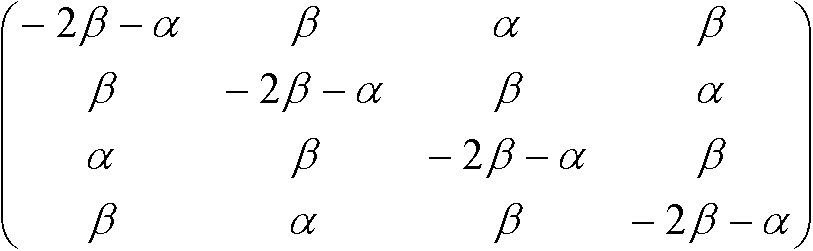 , ,置換行列: 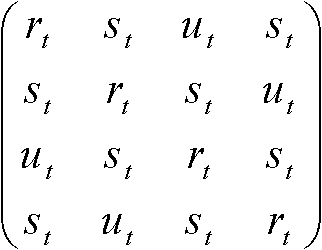
ただし、 最尤法進化系統樹の尤度, P(x*|T, t*) ≡ 配列集合x*が枝長t*の進化系統樹により生成される確率最尤法: 配列集合x*が与えられた時、尤度P(x*|T, t*)を最大化するT, t*を計算 残基の尤度: P(x1u, x2u|T, t1, t2) = ΣaqaP(x1u|a, t1)P(x2u|a, t2) 全体の尤度(配列長 = N): P(x1, x2|T, t1, t2) = Πu=1NP(x1u, x2u|T, t1, t2) Ex. C, Gのみから成る配列: n1個は同じで、n2個が変化 (Ex. n1 = 8, n2 = 3)
CCGGCCGCGCG
= 1/4(rt1rt2 + 3st1st2)
P(C, G|T, t1, t2) = 1/4(rt1st2 + 3st1rt2 + 2st1st2) = 1/16(1 – e-4α(t1+t2)) 系統樹全体の尤度, P(x1u, x2u, …, xnu|T, t*)
= Σqa2n-1Πi=n+12n-2P(ai|aα(i), ti)Πi=1nP(aui|aα(i), ti)
アルゴリズム: Felsensteinの尤度計算アルゴリズム P(Lk|a)) = Σb, cP(b|a, ti)P(Li|b)P(c|a, tj)P(Lj|c)
終了処理:
系統樹の形状探索 → 分枝限定法等 ブートストラップ法系統樹の信頼性: 推定された系統樹の信頼性不明 → 信頼性評価必要必ずしも系統樹全体でなく、大分岐等の特定特徴の信頼性評価で良い → ブートストラップ法による評価 ある確率モデルのもとでブートストラップ法による頻度 → 特徴の事後確率 系統樹に対するブートストラップ法
S(a, b) := –logP(b|a) ⇒ 最節約法のコスト ≈ 系統樹全体尤度の近似値(Viterbi近似)
葉以外の節点に対し
置換行列が乗法的かつ可逆的 |
|
新エングラー = Engler(-Prantl)体系 (Engler 1844-1930) 進化の経路: 単純 (雄蕊1/雌蕊1 = 原始的) ⇒ 複雑
双子葉 ⇒ 単子葉 |
多数雄蕊 ⇒ 少数雄蕊 多くの標本庫が、種子植物に関しては、この体系を使用している (Gray: not recorded from Hokkaido) |
| Cycadopsida ソテツ綱 | ||
|---|---|---|
Cycadales ソテツ目 |
Cycadaceae (ソテツ), Stangeriaceae, Zamiaceae (ザミア) |
Ginkgoales イチョウ目Ginkgoaceae (イチョウ) |
| Coniferopsida マツ綱 | ||
Coniferae (Pinophyta) マツ目Pinaceae (マツ), Taxodiaceae (スギ), |
Cupressaceae (ヒノキ), Podocarpaceae (マキ), Cephalotaxaceae (イヌガヤ), Araucariaceae (ナンヨウスギ) | Sciadopityaceae (コウヤマキ), not established |
| Taxopsida イチイ綱 | ||
Taxales イチイ目 |
Taxaceae (イチイ) | Chlamydospermae グネツム綱 |
Gnetales グネツム目 |
Gnetaceae, Ephedraceae (マオウ), Welwitschiaceae |
|
双子葉植物綱
古生花被植物亜綱 |
Archichlamydeae 古生花被植物亜綱Casuarinales モクマオウ目Casuarinaceae: no sp. in Jpn (exotic)Juglandales クルミ目Myricaceae (ヤマモモ), Juglandaceae (クルミ)Balanopales バラノプス目BalanopaceaeLeitneriales レイトネリア目Leitneriaceae, DidymelaceaeSalicales ヤナギ目Salicaceae (ヤナギ)Fagales ブナ目Betulaceae (カバノキ), Fagaceae (ブナ)Urticales イラクサ目Rhoipteleaceae, Ulmaceae (ニレ), Eucommiaceae (トチュウ), Moraceae (クワ), Urticaceae (イラクサ)Proteales ヤマモガシ目Proteaceae (ヤマモガシ)Santalales ビャクダン目Olacaceae (ボロボロノキ): Schoepfia Schreb. jasminodora Sieb. et Zucc.ボロボロノキ, 1 sp in Jpn (0 in Hokkaido), Dipentodontaceae, Opiliaceae (カナビキボク): 0 in Jpn, Grubbiaceae, Santalaceae (ビャクダン), Misodendraceae, Loranthaceae (ヤドリギ)Balanophorales ツチトリモチ目Balanophoraceae (ツチトリモチ)Medusandrales メドゥーサンドラ目MedusandraceaePolygonales タデ目Polygonaceae (タデ)Centrospermae アカザ目Didiereaceae (カナボウノキ), Phytolaccaceae (ヤマゴボウ), Gyrostemonaceae (ビャクブ), Achatocarpaceae, Nyctaginaceae (オシロイバナ), Molluginaceae (ザクロソウ), Aizoaceae (ハマミズナ), Portulacaceae (スベリヒユ), Basellaceae (ツルムラサキ), Caryophyllaceae, Dysphaniaceae, Chenopodiaceae (アカザ), Amaranthaceae (ヒユ)Cactales サボテン目Cactaceae (サボテン)Magnoliales モクレン目Ranunculales キンポウゲ目Ranunculaceae (キンポウゲ), Berberidaceae (メギ), Sargentodoxaceae (サルゲントドクサ), Lardizabalaceae (アケビ), Menispermaceae (ツヅラフジ), Nymphaeaceae (スイレン), Ceratophyllaceae (マツモ) |
Piperales コショウ目Saururaceae (ドクダミ), Piperaceae (コショウ), Chloranthaceae, LactoridaceaeAristolochiales ウマノスズクサ目Guttiferales オトギリソウ目Dilleniaceae (ビワモドキ), Paeoniaceae (ボタン), Crossosomataceae, Eucryphiaceae, Medusagynaceae, Actinidiaceae (マタタビ), Ochnaceae, Dioncophyllaceae, Strasburgeriaceae, Dipterocarpaceae (フタバガキ), Theaceae (ツバキ), Caryocaraceae, Marcgraviaceae, Quiinaceae (クイナ), Guttiferae (オトギリソウ), AncistrocladaceaeSarraceniales サラセニア目Sarraceniaceae (サラセニア), Nepenthaceae (ウツボカズラ), Droseraceae (モウセンゴケ)Papaverales ケシ目Papaveraceae (ケシ), Capparaceae (フウチョウソウ), Brassicaceae (Cruciferae, アブラナ), Tovariaceae, Resedaceae (モクセイソウ), MoringaceaeBatales バティス目BataceaeRosales バラ目Platanaceae (スズカケノキ), Hamamelidaceae (マンサク), Myrothamnaceae, Crassulaceae (ベンケイソウ), Cephalotaceae (フクロユキノシタ), Saxifragaceae (ユキノシタ), Brunelliaceae, Cunoniaceae, Davidsoniaceae, Pittosporaceae (トベラ), Byblidaceae, Roridulaceae, Bruniaceae, Rosaceae (バラ), Neuradaceae, Chrysobalanaceae, Connaraceae (マメモドキ), Fabaceae (マメ), KrameriaceaeHydrostachyales ヒドロスタキス目HydrostachyaceaePodostemales カワゴケソウ目Podostemaceae (カワゴケソウ)Geraniales フウロソウ目Limnanthaceae (リムナンテス), Oxalidaceae (カタバミ), Geraniaceae (フウロソウ), Tropaeolaceae (ノウゼンハレン), Zygophyllaceae (ハマビシ), Linaceae (アマ), Erythroxylaceae (コカノキ), Euphorbiaceae (トウダイグサ), Daphniphyllaceae (ユズリハ)Rutales ミカン目Rutaceae (ミカン), Cneoraceae, Simaroubaceae (ニガキ), Picrodendraceae, Burseraceae (カンラン),Meliaceae (センダン), Akaniaceae, Malpighiaceae (キントラノオ), Trigoniaceae, Vochysiaceae, Tremandraceae, Polygalaceae (ヒメハギ) |
Sapindales ムクロジ目Coriariaceae (ドクウツギ), Anacardiaceae, Aceraceae (カエデ), Bretschneideraceae, Sapindaceae (ムクロジ), Hippocastanaceae (トチノキ), Sabiaceae (アワブキ), Melianthaceae, Aextoxicaceae, Balsaminaceae (ツリフネソウ)Julianiales ジュリアニア目JulianiaceaeCelastrales ニシキギ目Rhamnales クロウメモドキ目Rhamnaceae (クロウメモドキ), Vitaceae (ブドウ), Leeaceae (ウドノキ)Malvales アオイ目Elaeocarpaceae (ホルトノキ), Sarcolaenaceae (サルコラエナ), Tiliaceae (シナノキ), Malvaceae (アオイ), Bombacaceae (パンヤ), Sterculiaceae (アオギリ), ScytopetalaceaeThymelaeales ジンチョウゲ目Geissolomataceae, Penaeaceaem, Dichapetalaceae (カイナンボク), Thymelaeaceae (ジンチョウゲ), Elaeagnaceae (グミ)Violales スミレ目Flacourtiaceae (イイギリ), Peridiscaceae (ペリディスクス), Violaceae (スミレ), Stachyuraceae (キブシ), Scyphostegiaceae (カミニンギョウ), Turneraceae (トゥルネラ), Malesherbiaceae (マレシェルビア), Passifloraceae (トケイソウ passion fruit), Achariaceae (アカリア), Cistaceae (ハンニチバナ), Bixaceae (ベニノキ), Sphaerosepalaceae (スファエロセパルム), Cochlospermaceae (ワタモドキ), Tamaricaceae (ギョリュウ), Frankeniaceae (フランケニア), Elatinaceae (ミゾハコベ), Caricaceae (パパイア papaya), Loasaceae (シレンゲ), Datiscaceae (ダティスカ), Begoniaceae (シュウカイドウ)Cucurbitales ウリ目Cucurbitaceae (ウリ)Myrtales (Myrtiflorae) フトモモ目Lythraceae (ミソハギ), Trapaceae (ヒシ), Crypteroniaceae, Myrtaceae (フトモモ), Dialypetalanthaceae, Sonneratiaceae (ハマザクロ), Punicaceae (ザクロ), Lecythidaceae (サガリバナ), Melastomataceae (ノボタン), Rhizophoraceae (ヒルギ), Combretaceae (シクンシ), Onagraceae (アカバナ), Oliniaceae, Haloragaceae (アリノトウグサ), Theligonaceae (ヤマトグサ), Hippuridaceae (スギナモ), CynomoriaceaeUmbelliflorae セリ目Alangiaceae (ウリノキ), Nyssaceae (ヌマミズキ), Davidiaceae (ハンカチノキ), Cornaceae (ミズキ), Garryaceae, Araliaceae (ウコギ), Apiaceae (セリ) |
Sympetalae 合弁花植物亜綱Diapensiales イワウメ目Diapensiaceae (イワウメ)
Ericales ツツジ目Clethraceae (リョウブ), Pyrolaceae (イチヤクソウ), Ericaceae (ツツジ), Empetraceae (ガンコウラン), EpacridaceaePrimulales サクラソウ目Theophrastaceae, Myrsinaceae (ヤブコウジ), Primulaceae (サクラソウ)Plumbaginales イソマツ目Plumbaginaceae (イソマツ)Ebenales カキノキ目Sapotaceae (アカテツ), Sarcospermataceae, Ebenaceae (カキノキ) 道植栽, Styracaceae, Lissocarpaceae, Symplocaceae (ハイノキ), HoplestigmataceaeOleales モクセイ目Oleaceae (モクセイ) |
Gentianales リンドウ目Loganiaceae (ホウライカズラ/マチン), Desfontainiaceae, Gentianaceae (リンドウ), Menyanthaceae (ミツガシワ), Apocynaceae (キョウチクトウ), Asclepiadaceae (ガガイモ), Rubiaceae (アカネ)Tubiflorae (Laminales) シソ目Polemoniaceae (ハナシノブ), Fouquieriaceae, Convolvulaceae (ヒルガオ), Hydrophyllaceae (ハゼリソウ), Boraginaceae (ムラサキ), Lennoaceae, Verbenaceae (クマツヅラ), Callitrichaceae (アワゴケ), Lamiaceae (Labiatae) (シソ), Nolanaceae, Solanaceae (ナス), Duckeodendraceae, Buddlejaceae (フジウツギ), Scrophulariaceae (ゴマノハグサ), Globulariaceae (ウルップソウ), Bignoniaceae (ノウゼンカズラ), Henriqueziaceae, Acanthaceae (キツネノマゴ), Pedaliaceae (ゴマ, tropical-subtropcial regions, not in Hokkaido), Martyniaceae (ツノゴマ), Gesneriaceae (イワタバコ, not in Hokkaido), Columelliaceae, Orobanchaceae (ハマウツボ), Lentibulariaceae (タヌキモ), Myoporaceae (ハマジンチョウ), Phrymaceae (ハエドクソウ) |
Plantaginales オオバコ目Plantaginaceae (オオバコ)Dipsacales マツムシソウ目Caprifoliaceae (スイカズラ), Adoxaceae (レンプクソウ), Valerianaceae (オミナエシ), Dipsacaceae (マツムシソウ)Campanulales キキョウ目Campanulaceae (キキョウ), Sphenocleaceae (ナガボノウルシ), Pentaphragmataceae (ユガミウチワ), Goodeniaceae (クサトベラ), Brunoniaceae, Stylidiaceae, Calyceraceae, Asteraceae (キク) |
Helobiales イバラモ目Alismatales when established subclass AlismatidaeNajadales イバラモ目Najadaceae (イバラモ), Potamogetonaceae (ヒルムシロ), Zannichelliaceae (イトクズモ) when separated from Potamogetonaceae (Ohwi merges them), Juncaginaceae (シバナ), Scheuchzeriaceae (ホロムイソウ)Hydrocharitales トチカガミ目Hydrocharitaceae (トチカガミ, one species in Hokkaido = Vallisneria L.セキショウモ (V. asiatica Miki セキショウモ)Alismatales(Alismales)/Helobiae オモダカ目/沼生目Alismataceae (オモダカ), Butomaceae (ハナイ), Limnochartitaceae (リムノカリス), Aponogetonaceae (レースソウ, cape-pondweed)Triuridales ホンゴウソウ目Triuridaceae (ホンゴンソウ) |
Liliiflorae (Lilianae), superorder ユリ上目Liliales ユリ目Liliaceae (ユリ), Xanthorrhoeaceae (ススキノキ), Stemonaceae (ビャクブ), Agavaceae (リュウゼツラン), Haemodoraceae (ハエモドルム), Cyanastraceae, Amaryllidaceae (ヒガンバナ), Hypoxidaceae (キンバイザサ)*, Velloziaceae, Taccaceae (タシロイモ), Dioscoreaceae (ヤマノイモ), Pontederiaceae (ミズアオイ), Iridaceae (アヤメ), Geosiridaceae, Burmanniaceae (ヒナノシャクジョウ), Corsiaceae, Philydraceae (タヌキアヤメ)* 大井 Amarylidaceae に合一 Juncales イグサ目Juncaceae, ThurniaceaeBromeliales パイナップル目Bromeliaceae (パイナップル)Commelinales ツユクサ目Commelinaceae (ツユクサ), Mayacaceae, Xyridaceae (トウエンソウ/キシリス), Rapateaceae, Eriocaulaceae (ホシクサ), Restionaceae (サンアソウ), Centrolepidaceae (カツマダソウ), Flagellariaceae (トウツルモドキ) |
Graminales イネ目Poaceae (Gramineae) (イネ)Principes ヤシ目Palmae (Palmaceae) (ヤシ)Synanthae パナマソウ目Cyclanthaceae (パナマソウ)Arales (Spathiflorae) サトイモ目Araceae (サトイモ), Lemnaceae (ウキクサ)Pandanales タコノキ目Pandanaceae (タコノキ), Sparganiaceae (ミクリ), Typhaceae (ガマ)Cyperales カヤツリグサ目Cyperaceae (カヤツリグサ)Zingiberales ショウガ目 (B-H, Scitamineae)Musaceae (バショウ), Zingiberaceae (ショウガ), Cannaceae (カンナ), Marantaceae (クズウコン), LowiaceaeOrchidales (B-H, Microspermae and Gynandrae) ラン目Orchidaceae (ラン) |
日本YList 植物名検索芽ばえ図鑑 北海道植物関連文献目録 日本生物多様性関連Webサイト一覧: リンク集 帰化植物・侵入植物侵入生物データベース [国立環境研究所] |
帰化植物一覧表 [岡山大学 野生植物研究室] 史前帰化植物 北海道外来種データベース 北海道自然環境研究センター北海道 エコネットワーク |
サケのふるさと館 厚岸水鳥観察館 夕張リゾート (西)オーストラリア「鉱山廃坑跡地における植物群集の再生」に関連Nature's Planet Museum: 藤原幸一 |