(Upload on September 26 2023) [ 日本語 | English ]
Mount Usu / Sarobetsu post-mined peatland
From left: Crater basin in 1986 and 2006. Cottongrass / Daylily
HOME > Plant taxonomy > Glossary > Fern (Pteridophyta)
Division Pteridophyta = ferns, sensu lato
The records of number of fern speciesLinne's Era: 11 gen. 190 spp.1793 Smith JE: 13 new genera → total = 24 genera Presence or absence of annulus (環帯) 1806 Schwarz: "Synopsis Filicum" → 36 gen. 700 spp1810 Willdenew: "Linne's Species Plantarum (5 edn)" → 41 gen. 1010 spp. 1961 Cronquist A:
Subkingdom II Embryophyta
Division 9 Bryophyta Life cycleRreproductive organ = sporangium(-gia) → sporeSpory (胞子型) a) Homosporous type (homospory) 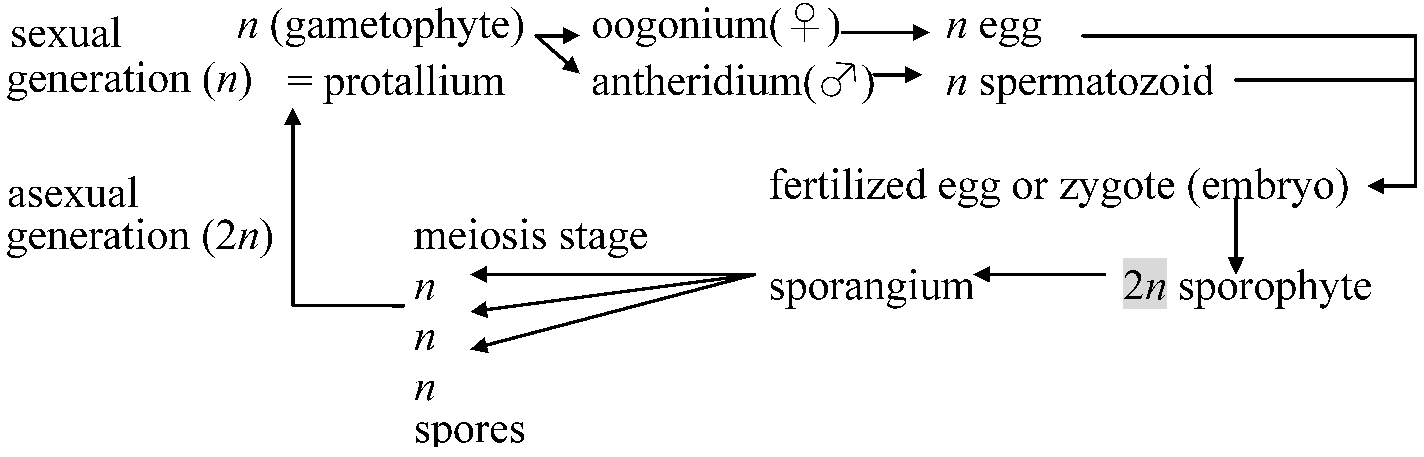 Ex. Pteridium aquilinum, Athyrium niponicum: divide primodal cell into two parts 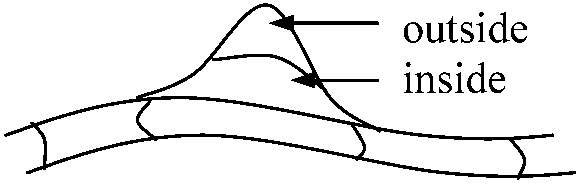 b) Heterosporous type (heterospory)
b) Heterosporous type (heterospory)microsporanium → microspore / macrosporanium → macrospore
┌─→ n (female gametophyte) → egg (n) ─────┐
│┌→ n (male gametophyte) ─→ spermatozoid (n)─┤
││ n ←─┐ 2n sporophyte
│└── n ←─┼─ microsporangium (2n) ←┐ │
│ n ←─┘ │ ↓
│ microspore ├ 2n sporophyte
│ n ←─┐ │
└─── n ←─┼─ macrosporangium (2n) ←┘
n ←─┘
macrospore
|
⇒ Engler's syllabus (エングラー体系), Gymnospermae, Angiospermae 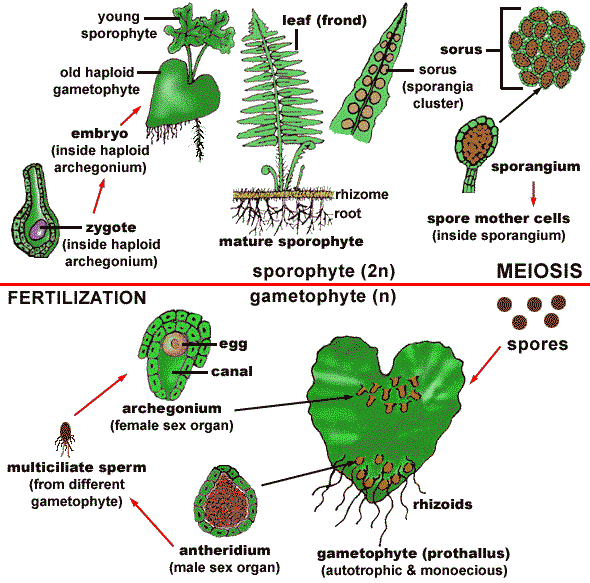 cp. Life cycle of moss
cp. Life cycle of moss
Pteridophyte coefficient (シダ係数)= pteridophyte indexrelated to climate (high air moisture and/or temperature) = (25 × number of fern species)/(number of seed plant species)Assumption: pteridophytes are sensitive to climatic factors, i.e., temperature, humidity and precipitation → Regions with high humidity and stable temperatures tend to have high pteridophyte richness |
[ morphology of seed plants]
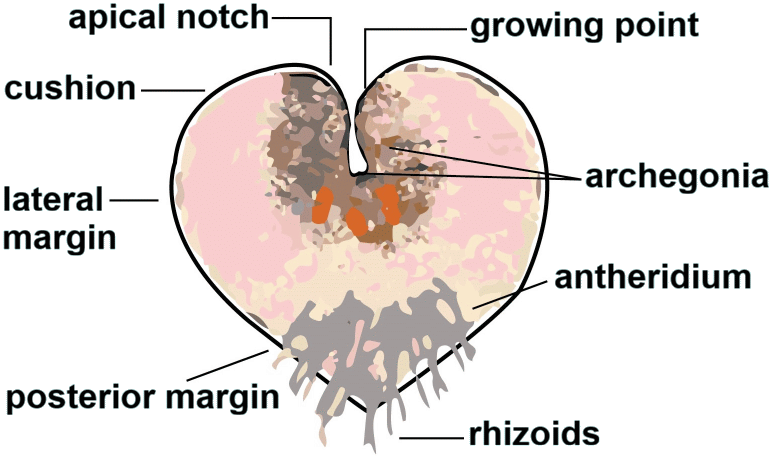
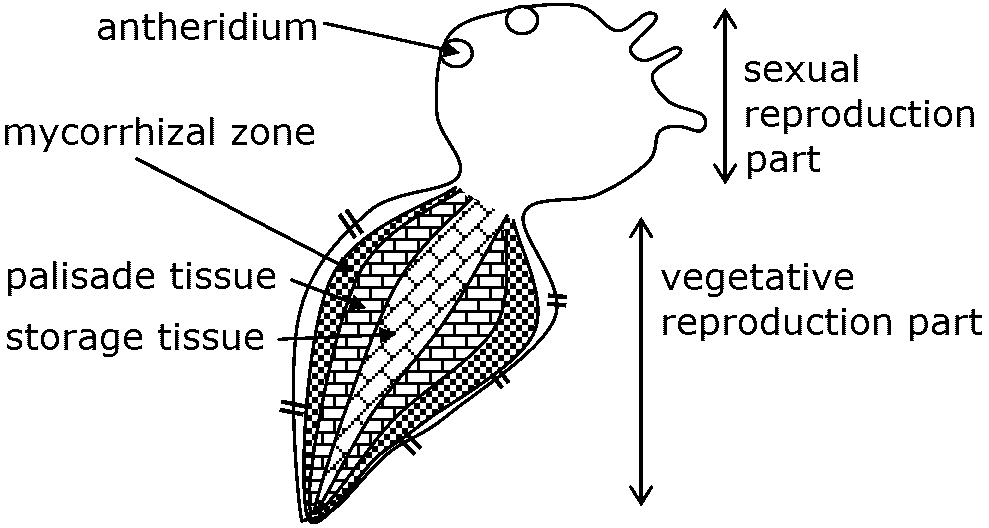
External morphology (外部形態)Prothallium (pl. -us. 前葉体) = gametophyte (配偶体)
increase in surface area to support water movement along the surface reducing water loss and protecting the growing tips in exposed or dry habitats Rhizoid (仮根): a slender, root‑like filament found on the prothallus
anchoring the gametophyte to the substrate and absorb water Appendage (付属物): any secondary structure attached to the main body Ex. scales, hairs Sporophyte (胞子体)

Frond (葉状体) ≈ leaf (葉) for fern, cydad and palm: The leaf of the fern bearing the pinnae = blade + stipe |
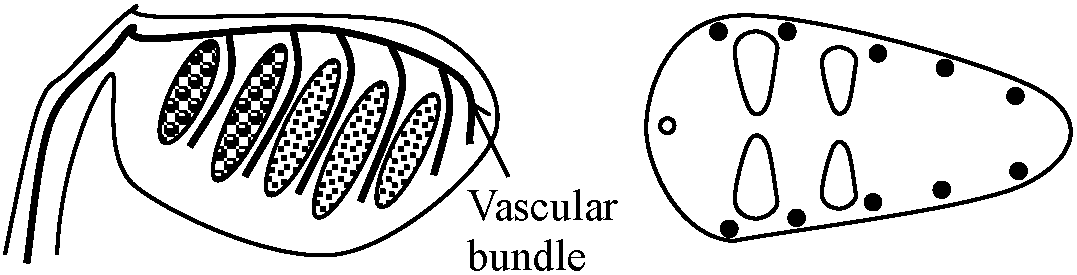
Blade (葉身): part of frond bearing the foliage Stipe (柄/茎) or petiole (葉柄): leaf stalk of the fern arising from the rhizome and below the blade Apex (頂端): the top of blade Midrib (中肋): central or principal vein of a frond Number of veins = the number of midrib branches of the fronds Pinnule(s) (小羽片) ≈ leaflet for seed plantsPinna (pl. -e) (羽片): the primary division of a compound fern frond. A distinct subdivision of the blade, when the blade segments are fully divided to the rachis Rachis (羽軸): the main stalk or central vein, of a fern frond that bears the pinnae - the blade section. The section below the pinnae is called the stipe 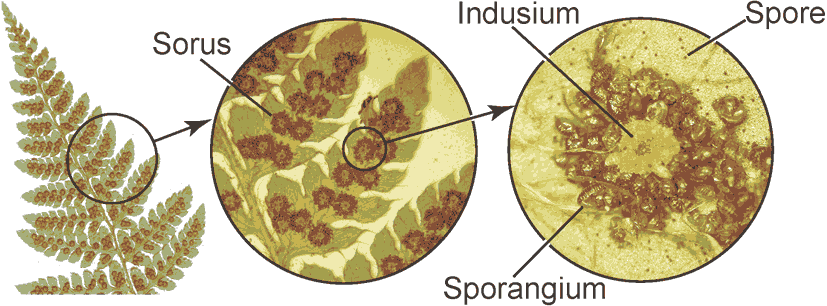 Sorus (pl. sori) (胞子嚢群): a cluster of sporangia arranged on the underside or margins of the pinnae in a distinctive pattern in each fern genus Indusium (pl. -a) (包膜): membranes covering the sori, forming a net-like or skirt-like shape Sporangium (pl. -a) (胞子嚢) for mosses and ferns: a spore case in which the spores develop. Thin walled, usually stalked Spore (胞子): a minute, typically one-celled, reproductive unit capable of giving rise to a new individual without sexual fusion Internal morphology (内部形態)Stele (中心柱)
|
[*: extinction]
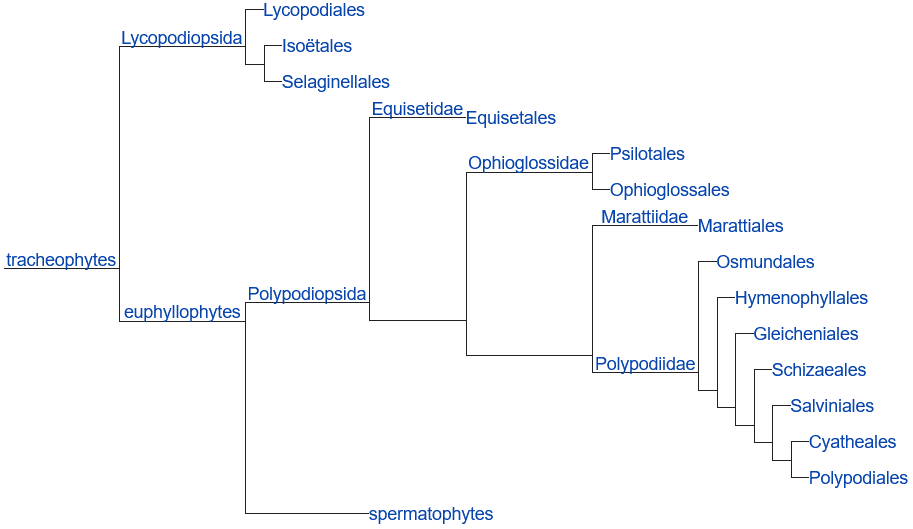 Phylogeny constructed by PPG I (2016) Rhyniopsida* リニアTrimerophytopsida* トリメロフイトンZosterophyllopsida* ゾステロフィルムPsilotopsida マツバランPsilotales マツバラン Lycopsida ヒカゲノカズラDrepanophycales* ドレパノフイクスProtolepidodendrales* 古生リンボク目(プロトレピデンドロン目) Lepidodendrales* リンボク(トレピデンドロン) Lycopodiales ヒカゲノカズラ |
Selaginellales イワヒバ Pleuromeiales プレウロメイア Isoetales ミズニラ Equisetopsida トクサ綱Hyeniales* ヒエニアPseudoborniales* プセウドボルニア Sphenophyllales* スフェノフィルム Equisetales トクサ目 Archaeocalamitaceae* Calamitaceae* Equisetaceae (トクサ) Phyllothecaceae* Filicopsida シダ綱Cladoxylales* クラドキシロンIridopteridales* イリドプテリス Rhacophytales* ラコフイトン Stauropteridales* スタウロプテリス Zygopteridales* ジゴプテリス Marattiales リュウビンタイ Ophioglossales ハナヤスリ Ophioglossaceae ハナヤスリ Filicales シダ Progymnospermopsida* 前裸子植物Aneurophytales* アネウロフイトンArchaeopteridales* アルカエオプテリス Protopityales* プロトピチス |
|
Dichotomy (叉状分枝) Protostele (原生中心柱) 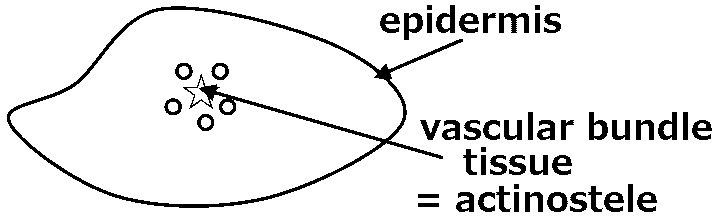 Terminal sporangium 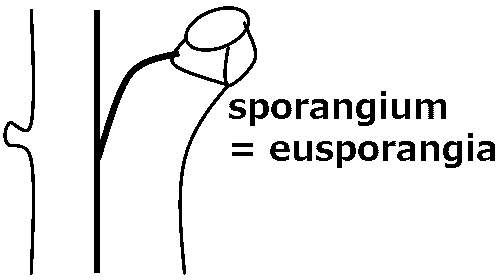 Class Psilophytopsida (古生マツバラン綱)*Silurian-DevonianClass Psilotopsida (マツバラン綱)The phylogenic relationship to Psilophytopsida is negativeExoscopic embryo Leafless and rootless Protostele  1-5: Psilotum triquetrum, 1 oberer Tell der Pflanze mit Sporangien, 2 geschlossenes Sporangium, 3 aufgesprung enes Sporangium, 4 Sporen, 5 Querschnitt durch ein noch unreifes Sporangium, 6 Spros, 6-7 Tmesipteris Sporophyll |
Order Psilotales (マツバラン)degeneration of microphyll → considered to be in microphyllous fern Psilotaceae (マツバラン)Psilotum L. P. nudum (L.) P. Beauv. (マツバラン) P. complanatum Sw. (ソウメングサ)  Fig. Psiltoum triquetrum (Gametophyte) Tmesipteridaceae (イヌナンカクラン), moved into Psilotaceae Tmesipteris Berhn., hanging fork fern |
|
= Division Lepidophyta (Lycopodophyta) (小葉植物植物門), clubmosses and quillworts 400-600 spp. in the world, 17 spp. in Japan, and 11 spp. in Hokkaido Spore: germination, growth
Gametophyta, e.g., Licopodium complanatum
palisade tissue (柵状組織)
The development of zygote suspensor (胚柄): non-divisive Class Lepidopsida (小葉植物綱)⇒ Aglossopsida (無舌綱) when Glossopsida is establishedOrder Protolepidodendrales (古生ヒカゲノカズラ*)DevonianOrder Lycopodiales (ヒカゲノカズラ)Carboniferous - presentLycopodium L. (ヒカゲノカズラ) Class Glossopsida (有舌綱)Order Lepidodendrales (リンボク目)*Devonian - PermianOrder LepidocarpalesThis order is discarded, when Lepidocarpaceae → Lepidodendales, Maidesmiaceae → SelaginellalesLepidocarpaceae*: Ledpidocarpon Scott Maidesmiaceae*: Miadesmia Betr. Order Isoetales ミズニラReproductive organs developed on the base of leaves in autumn (microphyllous) ↔ leaves produced in spring are vegetative organs only |
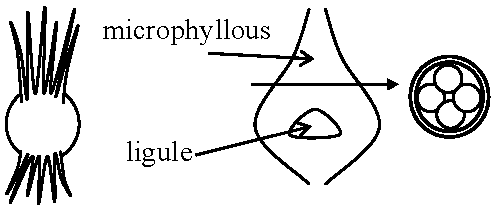 heterospory (異型胞子): macrospore – developing ealier, microspore – later eusporangiate (真嚢): germination
macrospore → free nucloeous division
Pleuromeia (Triassic): dioecious - related to Selaginellales (Mägdefran 1931, 43) Nathorstiana (Cretaceous) Stylites: heterosproy, fern-typed sperm (Rauh 1954) Isoetaceae (ミズニラ) Isoetes (ミズニラ): I. australis S.Williams (WA), I. drummondii A. Braun (WA), I. tripus A. Braun (WA) Stylites: discovered on the mountain of Pelu in 1954 Microphyllous → spiral microphyllous group = Isoetes / Stylites Order Selaginellales (イワヒバ)Carboniferous - presentSubterraniean gametophyte: mycotrophy (菌栄養) not developing chlorophyll Soprangium developing on the upper surface of sporophyll → phylogenetically different from Lycopodium 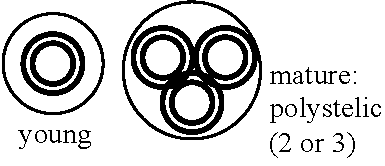 Differentiating leaves, stems and roots → Protostele Stem: slim = exarch (外原型) ↔ thick = endarch (内原型) Cf. Botryopteris (古生シダ) Root: rhizophore (担根体) by exogenous branching (or growth)
← roots are generally formed by endogenous branching
ligule (小舌): function unknown, absent in Lycopodiales |
Selaginella P. Beauv. (イワヒバ) in JapanS. biformis A. Br. ex Kuhn (ツルカタヒバ)S. boninensis Baker (ヒバゴケ) S. doederleinii Hieron. (オニクラマゴケ) var. opaca Seriz. (コウズシマクラマゴケ) S. helvetica (L.) Link (エゾノヒメクラマゴケ)S. heterostachys Baker (ヒメクラマゴケ) S. involvens(Sw.) Spring (カタヒバ), syn. S. gracillima (Kunze) Alston (Western Australia) |
S. leptophylla Baker (コケカタヒバ) S. limbata Alston (アマミクラマゴケ) S. lutchuensis Koidz. (ヒメムカデクラマゴケ) S. moellendorffii Hieron. (イヌカタヒバ) S. nipponica Franch. et Savat. (タチクラマゴケ) S. remotifolia Spring (クラマゴケ) S. selaginoides (L.) Link (コケスギラン) |
S. shakotanensis (Franch. ex Takeda) Miyabe et Kudo (ヒモカズラ) S. sibirica (Milde) Hieron. (エゾノヒモカズラ) S. tamariscina (Beauv.) Spring (イワヒバ), south to Oshima Peninsula and Okushiri Island S. tama-montana Serizawa (ヤマクラマゴケ) S. uncinata (Desv.) Spring (コンテリクラマゴケ) |
|
all of the present species are microphyllous type homospory (morophological): no suspensor, embryo developing outward
Sporophyll (= sporangiophore) Class Arthropsida (有節植物綱)horsetail and sconring rushesOrder Protoarticulales (Hyeniales)*Protohyeniaceae*Hyeniaceae*: Devonian: Hyenia (mid-Devonian) Calamophytaceae: identified by brancing patterns and separated from Equisetaceae formerly in the same group with Equisetaceae Order Sphenophyllales*Devonian -TriassicSpehno- [wedge] + phyllum [leaf] → sphenophyllous plant – herb-like Sphenophyllaceae
Protostele → siphonostele (管状中心柱) |
Order Cheirostrobales*CarboniferousCheirostrobaceae* Order Pseudoborniales*DevonianPseudoborniaceae* Order Calamitales (ロボク)*Devonian - TriassicSecondary auxetic growth - remarkable → stem: 30 cm φ, 20-30 m long Calamitaceae ロボク leaves primarily authorized as Annularia → later, combined with stem of which name is Calamites Calamites Suckow (ロボク): remarkable macrophyllous leaf Order Equisetales (トクサ)Carboniferous - presentEquisetaceae (トクサ) Equisetum L. (トクサ): only this genus is present now Chromosome number: n = 108 Herb-like = no secondary auxetic growth Tarminal sporangia = marginal Troll (1928), Göbel (1930) Strobus on Equisetum is "one" flower Scott (1912), Eames (1936), Ogura (1937)Counter by using Calamostachys and Paleostachya |
Equisetum L., horsetail (トクサ)E. arvense L. (スギナ)E. fluviatile L. (Syn. E. limosum L.) (ミズドクサ) E. hyemale L. (トクサ) E. palustre L. (イヌスギナ) E. pratense Ehrh. (ヤチスギナ) E. ramosissimum Desf. (イヌドクサ) |
var. ripense (Nakai et F. Maekawa) Tagawa (タカトクサ) E. scirpoides Michx. (ヒメドクサ)E. sylvaticum L. (フサスギナ) E. variegatum Schleich. ex Weber et Mohr (チシマヒメドクサ) |
Putative hybrids E. × moorei Newn. = E. ramosissimum × hyemale (トリトクサ) E. × rothmaleri C. N. Page = E. palustre × arvense (フォーリースギナ) E. × litorale Kuhlew. ex Rupr. = E. fluviatile × arvense (ハマスギナ) |
 Eusporangiate_______________________Leptosporangiate |
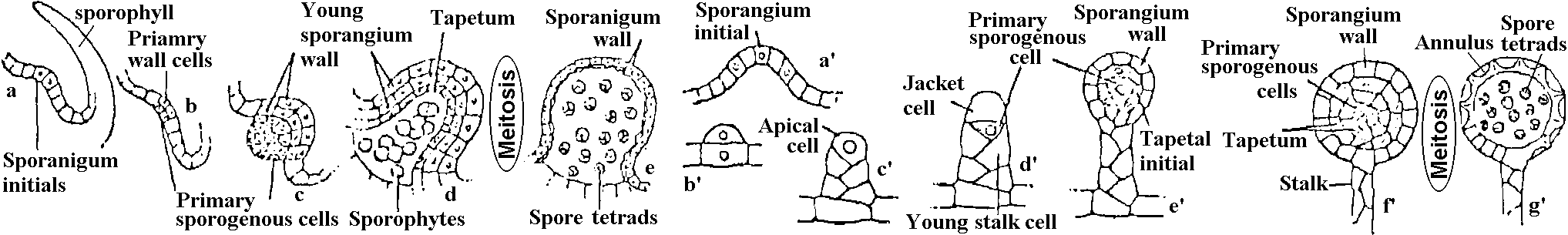
Fig. Ontogeny and structure of the two principal types of sporangia in vascular plants. a-e, the eusporangium; a'-g', the leptosporangium.
Subclass Primofilicidae (原シダ亜綱*)from mid-Devonian, flourish in CarboniferousSubclass Filiciedae 真正シダ亜綱a) Eusporangiate group (真嚢シダ類)= Ophioglossales + Marattiales
Ex. Ophioglossum, Botrychium b) Leptosporangiate (薄嚢シダ類)= Filicales (真正シダ目) + Marsileales + Salviniales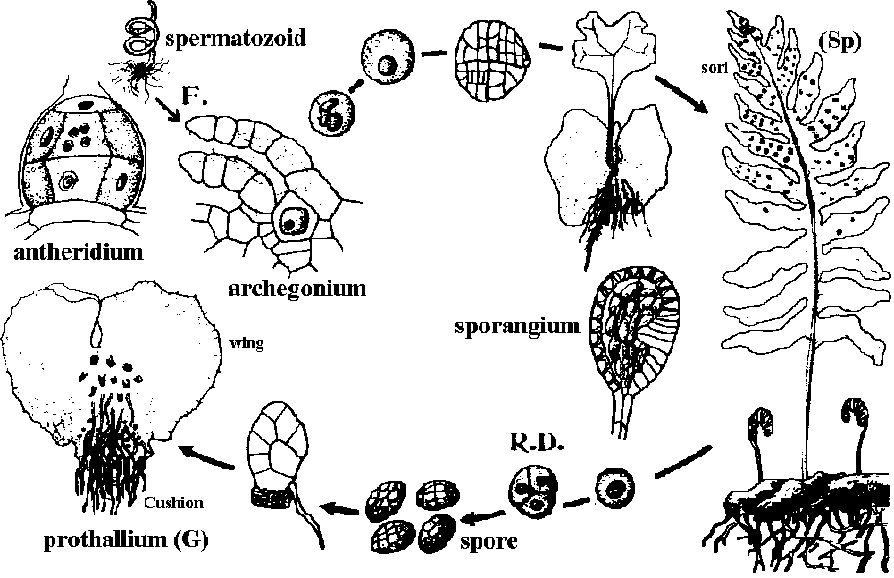 Fig. Life cycle of a fern, Polypodium vulgare (オオエゾデンダ) Order Eusporangiales (真嚢シダ目)Triassic-presentOphioglossaceae (ハナヤスリ): 4 gen. 70 spp. in the world Botrychium Sw. (ハナワラビ) in Japan B. atrovirens (Sahashi) M. Kato (シチトウハナワラビ) B. boreale (Fr.) Milde (タカネハナワラビ) B. daucifolium Wall. ex Hook. et Grev. (シマオオハナワラビ) B. formosanum Tagawa (ホウライハナワラビ) B. japonicum (Prantl) Underw. (オオハナワラビ) B. lanceolatum (S. G. Gmel.) Angstr. (ミヤマハナワラビ) ssp. angustisegmentum (Pease et A. H. Moore) Clausen (ホソバミヤマハナワラビ) B. lanuginosum Wall. ex Hook. et Grev. (アリサンハナワラビ)B. lunaria (L.) Sw. (ヒメハナワラビ) B. microphyllum (Sahashi) K. Iwats. (イブリハナワラビ) B. multifidum (S. G. Gmel.) Rupr. (ヤマハナワラビ) B. nipponicum Makino (アカハナワラビ) var. minus (H.Hara) K. Iwats. (ウスイハナワラビ) B. simplex E. Hitchc. var. tenebrosum (A. A. Eaton) Clausen (コケハナワラビ)B. strictum Underw. (ナガホノナツノハナワラビ) B. ternatum (Thunb.) Sw. (フユノハナワラビ) B. triangularifolium (Sahashi) M. Kato (ミドリハナワラビ) B. virginianum (L.) Sw. (ナツノハナワラビ) Hybrids B. japonicum (Prantl) Underw. × B. ternatum (Thunb.) Lyon (アイフユハナワラビ) B. × argutum (Sahashi) K.Iwats., nom. nud. (ハブハナワラビ) B. × elegans (Sahashi) K.Iwats., nom. nud. (アカネハナワラビ) B. × longistipitatum (Sahashi) K.Iwats., nom. nud. (アイイズハナワラビ) B. × pulchrum (Sahashi) K.Iwats., nom. nud. (アンコハナワラビ) Ophioglossum L. (ハナヤスリ) O. austroasiaticum M. Nishida (タイワンハナヤスリ) O. kawamurae Tagawa (サクラジマハナヤスリ) O. lusitanicum L. (WA) O. namegatae M. Nishida et Kurita (トネハナヤスリ) O. parvifolium Grev. et Hook. (イオウジマハナヤスリ) O. parvum M. Nishida et Kurita (チャボハナヤスリ) O. pendulum L. (コブラン) O. petiolatum Hook. (コヒロハハナヤスリ) O. thermale Kom. (ハマハナヤスリ) O. vulgatum L. (ヒロハハナヤスリ) O. × nipponicum Miyabe et Kudô (コハナヤスリ) Schizaeaceae Kaulf. (フサシダ) Schizaea Smith (S. fistulosa Labill.) Lygodiaceae C. Presl (カニクサ), when separated from Schizaeaceae Lygodium Sw. (カニクサ, climbing fern), one genus when this family is established: native to the tropical regions, consisting of ca 40 species Japan: L. circinnatum (Burm. f.) Sw. (ナンゴクカニクサ). L. flexuosum (L.) Sw. (シマカニクサ). L. japonicum (Thunb.) Sw. (カニクサ). L. microphyllum (Cav.) R. Br. (イリオモテシャミセンヅル), Yaeyama Islands Marattiaceae (リュウビンタイ)Angiopteris Hoffm. リュウビンタイ (A. lygodiifolia リュウビンタイ) Psaroniaceae (fossil species): Psaronius CottaOrder Protolepidosporangiales 前薄嚢シダ+ Osumudales ゼンマイ (included or separated)
Eusporangiagae → Seed plant (真嚢) Macrophyllous → morphologically-close to leptosproangiatae Stalk on sporangium becoming thick → sori developed only on sporophylls because of the specialization of sorus morphology Tapetum (絨毯細胞) revmoed and the outer layer remained → two-layered outer wall Homosporic: 300-500 spores ⇒ Formerly one genus, Osmunda, as described above After PPG I (2016), treated as 4-6 genera Osmunda, Osmundastrum, Claytosmunda, Plenasium, Leptopteris (NZ), Todea(Southern Hemisphere) Osmunda L. (ゼンマイ) in JapanO. banksiifolia (Pr.) Kuhn (シロヤマゼンマイ) O. cinnamomea L. (ヤマドリゼンマイ) O. claytoniana L. (オニゼンマイ) var. vestita (Wall.) Milde (タイワンオニゼンマイ) O. japonica Thunb. (ゼンマイ) |
O. lancea Thunb. (ヤシャゼンマイ): endemic to Japan, including southern Hokkaido O. nipponica Makino (コゼンマイ) var. divisa (Makino) Nakai (シシゼンマイ) O. × intermedia (Honda) Sugimoto (オオバヤシャゼンマイ)Metzgar JS, Skog JE, Zimmer EA & Pryer KM. 2008. The paraphyly of Osmunda is confirmed by phylogenetic analyses of seven plastid loci. Systematic Botany 33: 31-36 Order Leptosporangiales (薄嚢シダ)Carboniferous-present1954 German: 14 family, 5000-7000 spp. Loxomaceae Loxoma R. Br. ex A. Cunn., Loxomopsis Christ Pteridaceae Kirchn. (イノモトソウ) Sori on the leaf margin Forming coenosorus (thickened, roll-up inside) Rhizome = siphonostele Onychium Kaulfuss. (タチシノブ), sometimes separated from Pteridaceae by establishing Parkeriaceae Hook. (ホウライシダ) that is not allowed to use due to the rule of ICBN one species in Japan (10 species in the world) O. japonicum (Thunb.) Kunze (タチシノブ)
Pteridium Gled. ex Scop. (ワラビ) P. aquilinum (L.) Kuhn (ワラビ) Hypolepis Bernh. (コバノイシカグマ) Histiopteris J. Sm. (ヒメシダモドキ) Microlepia C. Presl (イシカグマ) Parkeriaceae or Adiantaceae (ホウライシダ) Adiantum L. (クジャクシダ) Anogramma Link: A. leptophylla (L.) Link, Western Australia Cheilanthes Sw., mostly in Western Australia: C. distans (R. Br.) Mett., C. sieberi Kunze, C. austrotenuifolia H. Quirk et T. C. Chambers Hymenophyllopsidaceae (コケシノブ) Hymenophyllum Smith (コウヤコケシノブ): H. barbatum (コウヤコケシノブ) Davalliaceae (シノブ) Plagiogyriaceae (キジノオシダ) Plagiogyria (Kunze) Mett. (キジノオシダ) (one genus in this family) Cyatheaceae (ヘゴ), merged into Aspidiaceae, s.l.) Sporocarp, sorus: Marsilea-typed emergence (as well as Dicksoniaceae
Cyathea Smith ヘゴ: C. fauriei ヘゴ、tree-like, evergreen Dicksonia, Cibotium, Culcita, Thyrsopteris Revised by Christenhusz et al. (2011) based on molecular phylogeny ⇒ Cibotiaceae (タカワラビ, s.s.): Cibotium Kaulf., separated from Dicksoniaceae Thelypteridaceae (ヒメシダ) Dryopteridaceae (s.s.), Aspidiaceae (s.l.) (オシダ) Arachniodes Blume (カナワラビ), holly fern (syn. Polystichopsis (J. Sm.) Holtt., カナワラビ) Dryopteris Adans. (オシダ) Polystichum Roth (イノデ): 200 spp. in world, 23 spp. in Japan Matteuccia Tadaro (クサソテツ) Onoclea L. (コウヤワラビ): two species in Japan
Nephrolepis cordifolia (L.) Presl (タマシダ) Blechoraceae (シシガシラ)Lindsaeaceae: Lindsaea Dryander ex Smith (L. linearis Sw., Western Australia) Aspleniaceae (チャセンシダ): > 650 spp. in 2 genera Asplenium L. (チャセンシダ): A. aethiopicum (Burm. f.) Bech., Western Australia Pleurosorus Fee: P. rutifolius (R. Br.) Fee, including Asplenium, Western Australia Hymenasplenium Hayata (ホウビシダ) Gleicheniaceae (ウラジロ), included into Aspidiaceae, s.l. Woodsiaceae イダデンダ, Athyrium is included in this family when established Athyriaceae (メシダ), included in Aspidiaceae, s.l. when established Matoniaceae (マトニア) Distribution: Malay Peninsula, Seram Island (Indonesia) Matonia R. Br.: M. pectinata R. Br., M. sarmentosa Baker Phanerosorus Copel.: P. sarmentosus (Baker) Copel., P. major Diels Polypodiaceae (ウラボシ) Vittariaceae (シシラン) Order HydropteridalesCretaceous or Triassic-present (sometimes, dividied into two orders, Marsileales/Salviniales)Common characteristics: sporocarp = sorus enveloped by indusium Different characteristics: Salviniales = float, annuals ⇔ Marsileales = submerged, perennials Order Marsileales (デンジソウ) when establishedMarsileaceae (デンジソウ): Marsilea L. (デンジソウ) → quadrifoliaOrder Salviniales (サンショウモ), when established Salviniaceae (サンショウモ) Azollaceaee (アカウキクサ): Azolla Lam. (アカウキクサ) A. caroliniana, A. imbricata (アカウキクサ), A. pinnata
|