(2025年3月22日更新) [ 日本語 | English ]
HOME > 植物リスト (Plant list) > ユリ科 (Liliaceae)
[ エングラー体系 | 単子葉植物 | 植物分類学 | 参考文献 ]
|
単子葉植物ユリ目(有里ではない) 目-科の区分は見解様々 多系群 ⇒ APG分類体系(科レベルで大幅変更) 200 gen. 3000 spp (s.l., or Engler) 花: 両性(稀単生)、多く放射相称 花被弁: 外3弁 + 内3弁 = 6弁 (2列), 離生-合生 ⇒ 離生花被 apotepalous: 萼と花弁が分化しない花被で離弁に相当する形態 雄蕊: 6 → 葯縦裂普通 子房: (多くは)上位, 3(稀1室), 3個の側膜胎座
生活型: 普通多年生草本(稀蔓生低木)
クロンキスト エングラー体系Liliaceae (ユリ)Xanthorrhoeaceae (ススキノキ) Stemonaceae (ビャクブ) Agavaceae (リュウゼツラン) Haemodoraceae Cyanastraceae Amaryllidaceae (ヒガンバナ) Hypoxidaceae (キンバイザサ) Velloziaceae Taccaceae (タシロイモ) Dioscoreaceae (ヤマノイモ) Pontederiaceae (ミズアオイ) Iridaceae (アヤメ) Geosiridaceae Burmanniaceae (ヒナノシャクジョウ) Corsiaceae Philydraceae (タヌキアヤメ) |
ダルグレン APGCorsiaceae (コルシア)Campynemataceae (カンピネマ) Melanthiaceae (シュロソウ) Petermanniaceae (ペテルマニア) Colchicaceae (イヌサフラン) Alstroemeriaceae (Luzuriagaceae含) (ユリズイセン) Rhipogonaceae (リポゴヌム) Philesiaceae (フィレシア) Smilacaceae (サルトリイバラ) Liliaceae (ユリ) 科の検索(日本)1. 子房下位 ovary inferior2. 雄蕊6 (6 stamens)、葯外向きでない。葉跨状でない 3. 花両性。茎蔓にならない
4 葉分裂しない。子房3室 ___ Amaryllidaceae ヒガンバナ 2 雄蕊3 (3 stamens)、葯は外向き。葉は跨状 ___ Iridaceae アヤメ 1 子房上位 ovary superior
2 花被片鱗片状 ___ Juncaceae イグサ
3 花3数性trimerous (稀2数性)。子房3-2室 ___ Liliaceae + Asparagales キジカクシ(アスパラガス) + Xanthorrhoeales ススキノキ |
(北村他 1964)改
|
Subfam Melanthoideae シュロソウ (Melanthiaceaeとし独立)
Chionographis Maxim. シライトソウ Herreriopsis H. Perrir: マダガスカル Subfam Asphodeloideae ツルボラン (☛ 独立 Asphodelaceae)
Aloe L. アロエ Subfam Lilliodeae ユリ
Amana Honda アマナ Subfam Scilloideae ツルボ (Scillaceaeとし独立) Scilla ツルボ Subfam Allioideae ネギ Subfam Asparagioideae キジカクシ
Asparagus L.キジカクシ |
Subfam Ophiopogonoideae ジャノヒゲ (Mondoideae) (Ophiopogonaceaeとし独立させる見解)
Ophiopogon Ker-Gawl. ジャノヒゲ Cf. 亜科位置調べる: Narthecium asiaticum キンコウカ - 道EN APG: ユリ科から独立 (🔽APG IV) Asphodelaceae Juss. ツルボランSubfam Asphodeloideae (ツルボラン)Aloiampelos, Aloidendron, Aristaloe, Asphodeline, Asphodelus (ツルボラン), Astroloba, Bulbine, Bulbinella, Chortolirion, Eremurus, Gasteria, Gonialoe, Haworthia, Haworthiopsis, Jodrellia, Kniphofia (シャグマユリ), Kumara, Trachyandra, Tulista Aloe L. (アロエ) A. vera (L.) Burm. f. (バルバドスアロエ): アロエという時は本種のことが多い 🍹葉(若-古まで) - 熟成2ヶ月 - 中身そのまま
アロイン、アロエエモジン、粘質素、Vit B, C群、苦味質 

生活型: 多年生多肉植物 分布: 南アフリカ、ケープ州。オーストラリアに定着 Subfam Hemerocallidoideae (キスゲ) Pasithea, Phormium (マオラン), Agrostocrinum, Geitonoplesium, Thelionema, Rhuacophila, Herpolirion, Stypandra, Eccremis, Dianella (キキョウラン), Simethis, Hemerocallis (ワスレグサ), Tricoryne, Corynotheca, Caesia, Hodgsoniola, Arnocrinum, Stawellia, Hensmania, Johnsonia Subfam Xanthorrhoeoideae (ススキノキ) Xanthorrhoea (ススキノキ) |
|
P. odoratum (Mill.) Druce (アマドコロ, s.s.) var. maximowiczii (Fr. Schm.) Koidz. (オオアマドコロ) 5-6月に葉の付根から白花を1-2個づつ付ける 4-5月未開葉の筍状の部分 ☛ 山菜 全草に甘味 = 名前 (癖なくうまい + 大きくても食べられるが筋堅い) |
🧑🍳 (下処理) 茹でて苦味なくなるまで水に晒す →
天ぷら、油炒め、煮物、卵とじ、お浸し、酢味噌和え、酢の物 |
|
7(8)種 1 花萼片のみ。萼片鈍頭、長さ12-20 mm _____ エンレイソウ 1 花萼片と花弁。萼片長20-35 mm。葯6-15 mm。子房円錐状卵形
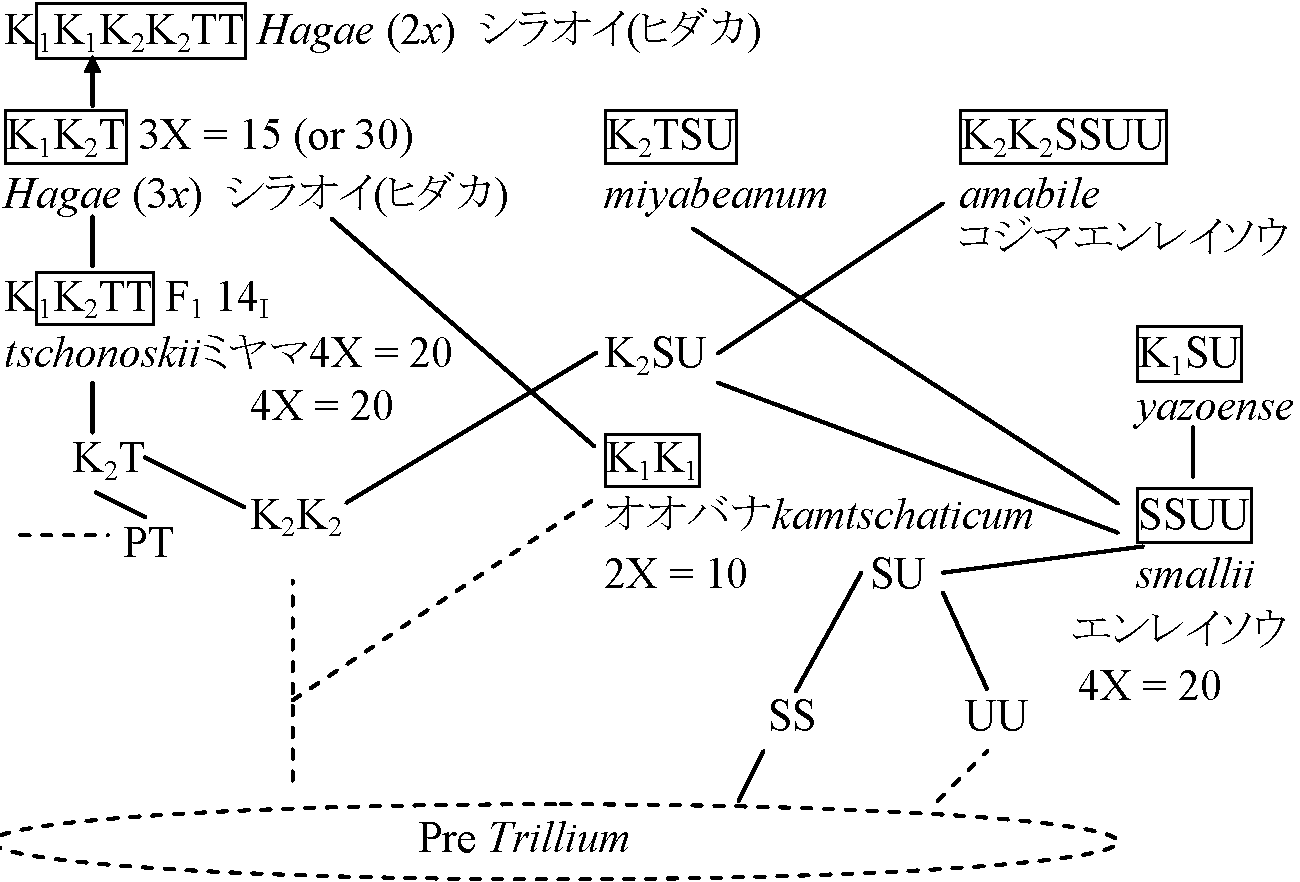
2 萼片花弁鋭頭12-27 mm l。葯長さ6-7 mm。花柄2-3 cm _____ シロバナエンレイソウ(= ミヤマエンレイソウ) T. smallii Maxim.(non. T. apetalon Makino) エンレイソウ: 地下茎太く地下這う丈夫な根。茎: 高さ20-40 cm, 円く無毛 T. amabile Miyabe et Tatewaki コジマエンレイソウ = エンレイソウ × オオバナ: エンレイソウに似るが紫色花弁-より発達。花弁広卵形 T. × miyabeanum Tatewaki ヒダカエンレイソウ = シロバナ × エンレイソウ: コジマに比べ萼片先端鋭い。子房卵形。雄蕊は雌蕊と同長か長い T. × hagae Miyabe et Tatewaki シラオイエンレイソウ = シロバナ × オオバナ: 花が横を向く、大型(高さ20-50 cm)染色体遺伝学染色体基本数= 5 ⇒ 全て倍数関係実験: 染色体低温処理 - Feulgen negative region - ゲノム分析 - 類縁関係は次の様に明らか (Kurabayashi & Saho 1957)
|
|
|
E. japonicum Dence. (カタクリ): 日本1種 - 万葉集に見る(堅香子)
☛ 山菜: 4-5月 葉・茎・花 + 地上部枯れた頃 鱗茎(乱獲注意)
過去には片栗粉原料 - 奈良県で栽培された |
🧑🍳 (下処理) 熱湯を潜らせる程度
お浸し、和え物、酢醤油、煮物、煮浸し、油炒め、汁の実、天ぷら |
|
APG: Asphodelaceae (ワスレグサ) 若芽、若葉、蕾・花 ☛ 山菜(蕾・花, 金針菜)
北海道: エゾカンゾウ(代表) - 本属の種は可食 (四川ではよく食べた) var. fluva (ホンカンゾウ) var. kwanso Regel (ヤブカンゾウ) var. littorea (Makino) M. Hotta (syn. H. littorea Makino) (ハマカンゾウ) var. longituba (Miq.) Maxim. (syn. H. longituba Miq.) (ノカンゾウ) |
🧑🍳 (下処理) 生(気になるなら茹でる) →
生(サラダ)、天ぷら、炒め物、ぬた (保存) 蕾・花乾燥 (やらん)
アスパラギン、アルギニン、ペントザン、アデニン、コリン、ブドウ糖、タンパク質 |
|
G. lutea (L.) Ker-Gawl. (キバナノアマナ) 春開花期 葉・茎・花食用 ☛ 山菜 |
鱗茎は一年中(ただし地上部が枯れた以降がよく成長している) 🧑🍳 (下処理) 熱湯を潜らせる(それ以上しない) →
生、汁の実、天ぷら、お浸し、油炒め、和え物、煮付、三杯酢、酢醤油 |
|
1. 花 > 10 cm l。夜間に開花し芳香ある。雄花は花被基部に合生 ___ H. plantaginea (ギボウシ) 1. 花 < 5 cm l。朝か昼間に開花し芳香ない。雄蕊は花被基部と離生 2. 葉狭く長楕円状披針形-長楕円状卵形、基部は葉柄に流れる。花茎直立し苞舟形、花冠多くは濃紫色
3. 花茎には普通の葉が多くつき苞も葉状、花柄短く8-12 mm ___ H. undulata (スジギボウシ) 4. 苞舟形で、蕾時は重なり開花の時も立つ
5. 葉縁は黄色、葉は根際から水平に開出する ___ ニシキギボウシ 6. 花冠の筒狭部は筒広部より短い
7. 葉鋭頭-鋭尖頭。葉身は葉柄に次第に移行する ___ H. rectifolia (タチギボウシ) 6. 花冠の筒狭部は筒広部と同長
7. 葉狭披針形で立ち、光沢があり、花小さく少ない ___ ミズギボウシ 8. 葉は水平に開出。苞は自身で巻く ___ ハカマギボウシ
9. 全体大、花茎80-120 cm、葉30-40 cm ___ オモトギボウシ 4. 苞は扁平で、蕾の時は開出し星状となる 5. 全体大、花茎45-70 cm、葉30-45 cm
6. 葉狭卵形、苞残る ___ ナンカイギボウシ 5. 全体より小、花茎20-30 cm、葉10-15 cm
6. 花被片披針形 ___ ヒメイワギボウシ 2. 葉広く卵形-広卵形、基部は心形-円形または切形、葉柄に流れない(稀に狭く流れる)
3. 花冠筒広部ほぼ直角に広がりコップ状となる ___ ムラサキギボウシ 4. 苞は舟形、広楕円形、2 cm l、やや厚い。花茎に筋がある。花穂は短く花が集まる |
5. 葉1-2枚、葉身20-40 cm、裏面緑白、花間やや離れる ___ ウラジロギボウシ 4. 苞扁平、花筒狭部に溝なく、筒広部透明線は不明
5. 苞0.7-1.2 cm、白紫色で薄膜質、花後に腐る。花9-10月、葉9-13 cm ___ イワギボウシ 6. 苞最下片長く長鋭尖頭、その2片は蕾時内に花序を抱き円錐体作る
7. 花茎は下へ曲がって垂れる ___ ウナズキギボウシ 6. 苞短く覆瓦様になる
7. 筒広部急に広がり、花冠裂片反り返る ___ ラッパギボウシ 6. 苞短く扁平で、蕾時星状に展開し、しばしば帯白色。花被は質厚い
7. 葉質硬く厚く平たいか舟状、初め白粉ある。花茎高く、花糸太い。下面側脈は隆起せず脈状突起ない ___ ラッパギボウシ 8. 花茎は葉叢より短いか少し高い。葉は碧緑白色、筒の広部は円錐形
9. 葉大きく葉身20-25 cm、広楕円形。栽培 ___ H. sieboldiana (トウギボウシ) 8. 花茎は葉叢より著しく長い。葉緑色。筒広部狭円錐形
9. 葉身15-30 cmで有長柄 ___ H. sieboldiana (オオバギボウシ) ギボウシ類: 若い葉柄や葉身はウルイと呼び食用 ☛ 山菜 🧑🍳 (下処理) 葉柄(開葉前は葉も可)、苦味あれば茹でる →
天ぷら(蕾・花)、汁の実、お浸し、和え物、三倍酢 オオバギボウシ: 山地等のやや湿った草地に生える大型草本
古くから観賞用に植えられ園芸品種多 |
|
1. 茎草質で刺ない。葉とともに冬は枯れる
2. 葉裏面が淡緑色で光沢。花7-8月に開き葯は線形で1.5 mm l ___ シオデ S. riparia 2. 液果は熟して赤色。茎に刺
3. 散形花序には多数の花がつく。葉大型で巻き髭は長い ___ サルトリイバラ S. china 2. 液果は熟して黒色か紫黒色。茎に刺のあるものとないものがある 3. 刺は殆どないか全くない。葉裏面は白色を帯びる |
4. 巻き髭がない。花序は2-5花からなる ___ マルバサンキライ
3. 刺は多いか疎ら。葉裏面は白色を帯びない。巻き髭があり、花序は多数花からなる
4. 刺は多い。花軸は分枝せず、花被片は淡黄緑色で開花時に反り返らない ___ ヤマカシュウ シオデ S. riparia 6-7月 太い若葉(柔らかければ可食) ☛ 山菜 (タチシオデも同様) 🧑🍳 (下処理) 採ったら早く茹でる(でないと黒変) →
天ぷら、汁の実、煮物、お浸し、和え物、バター炒め |
APG III: Asparagaceae
|
O. jaburan (Siebold) Lodd. (ノシラン) O. japonicus (Thunb.) Ker Gawl. (ジャノヒゲ) テンモンドウ(天門冬) = 生薬名 🍹秋-冬に長狭球形部分掘る - 洗って数分熱湯に浸し天日乾燥後に貯蔵 |
熟成3ヶ月 - 中身そのまま |
[APG: Colchicaceae]
|
🍹蕾・花 - 熟成2ヶ月 - 中身引抜く
マンノーズ、ガラクトーズ、ペントーズ、アラビノーズ、ブドウ糖 有毒成分: サポニン saponins - 消化器症状(嘔吐・下痢・腹痛) |
サポニンはアルコールに可溶 - 山菜・山野草酒に使わない D. sessile D. Don ex Schult. et Schult. f. (ホウチャクソウ) アマドコロ似 ⇒ 区別点: 葉のツヤ。ホウチャクソウ: 上部で枝分かれ多く根はヒゲ根のみ ↔ オオアマドコロ: 枝分かれなく地下茎伸びる |
|
1. 外花被片白色, n = 8。葉 20-30 cm l。地下茎は太い ___ キヌガサソウ P. japonica 1. 外花被片緑色。葉 ≈ 10 cm l |
2. 葉は普通8。内花被片糸状。葯壁長く伸びる ___ クルマバツクバネソウ P. verticillata |
|
= ニシキユリ(錦百合) 短日植物 地中海東部沿岸原産: 1863年に日本移入 花: 野生種 = 青紫色 - 園芸種 = 多様(青紫、白、ピンク、赤、橙、黄色他) |
蒴果: 卵円形 鱗茎: 卵形 H. orientalis var. albulus (ローマンヒアシンス): 疎らな小さめの花 |
[APG III: キジカクシ/クサスギカズラ科]
|
世界3種 日本2種 M. dilatatum (マイヅルソウ)
葉: 2葉卵心形、微凸頭/鋭頭3-12 cm |
M. bifolium (L.) F. W. Schmidt (ヒメマイヅルソウ): より小型
葉: 微細な鋸歯(ルーペ必要) |
|
ユキザサのみ道分布 S. japonica A. Gray (ユキザサ, 雪笹) 4-5月 新芽・若葉 (結構大きくなってもいける) ☛ 山菜 🧑🍳 (下処理) 不要 → |
嫌みなく甘味あり柔らかい。持ち味を生かした料理をして食べたい S. robusta Makino et Honda (ハルナユキザサ, 榛名雪笹) S. viridiflora Nakai (Syn. S. hondoensis Ohwi) (ヤマトユキザサ, 大和雪笹) |
北半球96種園芸品種Asiatic hybrid (Lilium × elegans) 親は主に橙色カロチノイド色素持つ: アジア原産親(L. dauricum, L. maculatum, L. callosum, L. pumilum, L. cernuum, L. lancifolium 等)を中心に交配 Oriental hybrid 日本産ユリ: 親 = L. auratum, L. speciosum、タモトユリ等 - 森林性 Longiflorum hybrid テッポウユリ群: 親 = L. formosanum, L. longiflorum Martagon hybrid (マルタゴン): 親 = L. martagon, L. hansonii, L. medeoloides等 Trumpet hybrid: 親 = 中国原産 L. henryi, L. regale (regal lily), L. brownii var. viridulum等 その他食用ユリ (作物)大半を北海道で生産
アイヌが珠芽を食用とした |
オオウバユリ L. cordatum var. glehnii
姥百合語源: 花の咲く頃には葉(歯)がない
和え物、味噌煮、天ぷら 
 Table Dance 
 Tiny Goast |

 Lollipop 
 Ceb Dazzle Notes ヤマユリ: 夏に香りの強い大きな花が咲く。花は白色地に斑点と黄色い線がある。山野の傾斜地や草叢に自生する。鱗茎は百合根と呼び古くから食用 |
日本の固有種は海を渡るEndemic Japanese Species Cross the Oceansユリはユリ科(Liliaceae)ユリ属(Lilium)の球根植物で、北半球の亜熱帯から亜寒帯にかけて自生している。日本にはこのうち15種が自生し、その半数は日本の固有種である。球根を食用とし、救荒植物として扱われてきたほか日本最古の書物「古事記」(712年)や「万葉集」(806年頃)にも登場し、古来より日本人にとって身近な花であった。近年では自生種の数が減少している。 日本での品種改良の歴史は古く、江戸時代には著名な植木屋伊藤伊兵衛の「花壇地錦抄」によりスカシユリ等(Lilium maculatum cv.)の園芸品種が紹介され、江戸園芸文化の一翼を担っていたと思われる。17世紀以降、プラントハンターの活躍が盛んになり、Kaempfer, Sieboldがユリの球根を欧州に持ち帰った。多花性であるが花自体は小さい欧州のユリと比べ、大輪で野性味がある日本のユリが欧州で注目を浴びるようになった。 |
明治期になり球根の輸出が始まると急激にその輸出額が伸び、日本にとって重要な輸出品目の1つとなった。ヨーロッパに渡った日本のユリは品種改良され、数多くの園芸品種が生まれた。現在、世界中で楽しまれているユリの多くは、日本に自生しているユリの血が脈々と流れ、素晴らしい園芸品種がオランダ、ニュージーランド等から日本に逆輸入されている。

 日比谷公園サービスセンター (2025/04/07) |
(大学3年時 実験ノート)
ユリ科植物
コルヒチン colchicineC22H25NO6, m.w. = 399.44 g/mol: 天然(トロポロン)アルカロイド - 三環構造
Colchicum autumnaleから抽出 - 二次代謝産物 生物学実験利用微小管形成阻害作用: チュブリン tublin に結合しチュブリンの重合を阻害 → |
微小管の形成・維持阻害 ⇒ 細胞周期研究: 細胞周期制御: 細胞分裂中期に停止 → 染色体観察容易 ∵ 微小管は染色体分離に関与 ⇒ 倍数体研究: コルヒチン → 細胞分裂中断 →細胞分裂未完了のままDNA複製 → 染色体数倍化 薬効作用機序: 白血球(特に好中球)微小管形成を阻害 → 炎症反応抑制痛風発作緩和・予防、家族性地中海熱(遺伝性疾患)緩和 副作用: 胃腸障害(下痢、腹痛、吐気)
+ 稀に再生不良性貧血や横紋筋融解症等重篤な副作用 |
花粉母細胞減数分裂⇒ meiosis of pollen mother cells減数分裂中期染色体構造と金属イオン減数分裂の中期染色体構造に及ぼす金属イオンの影響 (R.D. of pollen mother cells on Trillium)材料と方法 (これまでソラマメ(Vicia faba)根端細胞観察実験に基づき体細胞分裂過程を把握) ついで、減数分裂過程を理解する。また、その過程中に金属イオンを投与しその結果を考察する 準備: メスシリンダー・ピペット・ビーカー・メス・スライドグラス・カバーグラス・ピンセット・割箸・ロウ・へら・スパチュラ・濾紙・酢酸カーミン・Kcl・NaCl・CaCl2・KNO3 手順
|
X線による細胞分裂異常: 特に染色体異常誘発目的: Trillium属の核型を観察しkaryotypeを決定する。ゲノム解析genome analysisの基礎を知る。材料にX線照射を行い予想される染色体異常を観察する。染色体構造変化を目的としたX線を用いた染色体異常誘発を観察するI. X線(電離放射線)による異常細胞分裂異常
spindle fiber異常 → 遅滞染色体lagging chromosomes 体細胞ではG1-S期照射は染色体異常、S-G2-M期照射は染色分体異常、M期前期照射は半染色分体異常が生じた。減数分裂では第1分裂前期太糸期-中期照射時に半染色分体異常を生ずるのが特異的である II. 材料と方法材料: Trillium kamtschaticum Pallの胚珠壁細胞(ovular tissure cells)方法: ovaryにx1 r, x2 r, x3 rのX線照射 (x r/min; Kv, mA; mm Cu + mm Al) = ここでは20R, 40R, 80R 固定・染色: 照射後、24h, 48h, 72hで固定する
Fixation (LaCour 2BE fixative使用なら20 min.) III. 観察
|
|
Trillium染色体の euchromatic and heterochromatic segments 観察 1. 目的: 中期染色体を色素染色するとDNA合成時期のずれから、ある条件下で濃染部分(= 真正染色質euchromatin)と非濃染部分(= 異質染色質heterochromatin)を識別できる。唾液染色体では縞部分bandはeuchromatin、縞-縞間不染色部分やクロモセンターはheterochromatinである。エンレイソウを低温状態に長時間おくと中期染色体はFeulgen染色非染色部分が見られ、この様なFeulgen stain negative segmentによる縞模様は個体特異性がありエンレイソウ集団遺伝学的解析に利用される |
2. 方法: オオバナノエンレイソウ: 0°Cで72時間放置後、ovaryを前実験(X線染色体異常)の要領で固定染色しプレパラート作製。固定液はice bathで冷やし固定中も冷たい条件で行う 3. 観察: 退色模様の出ている中期染色体をスケッチする |
Trilliumにみられる倍数体の染色体観察方法: Trillium数種のovular tissue cells染色体を"X線染色体異常"の項の方法で固定染色しsquash methodでプレパラート作製準備: ビーカ・メスシリンダ・細口試薬ビン・冷却器・分液漏斗・温度計・ピペット・濾紙・薬包紙・45% acetic acid・basic fuchsin・1N HCl・無水異性重亜硫酸カリ・活性炭・1% クロム酸カリ・5% acetic acid・サポニン・2% osmic acid・恒温槽・管ビン・ガーゼ・カミソリ・輪ゴム・slide glass・ピンセット・割箸・ロウ・ヘラ 材料: Trillium (T. apetalon, T. tschonoskii, T. hagae, T. miyabenum, T. kamtschaticum) - these ovaries 操作A. Observation of karyotype
|
シッフ試薬(Schiff's reagent)作成法アルデヒド検出試薬 (= フクシンアルデヒド試薬)シッフ反応 (Schiff’s reaction): アルデヒド存在下で2分子アルデヒドR(O=)CHと1分子Schiff's fluidが反応し赤紫色化合物生成 Basic fuchsin (1 g) + 200 ml boiled water↓ 良く溶かす ↓ 50°Cまで冷やす 20 mlの1N HClを加える [= 1N HCl: specifi gravity = 1.19のconc. HCl 82.5 mlを水で1 lにする] ↓ 25°Cまで冷やす ↓ 無水異性重亜硫酸カリ3 gを加える(別名ピロ亜硫酸カリK2S2O5) [← 無水亜硫酸ソーダNa2SO3代用可] 冷暗所に24hr以上置く(黄色となる) |

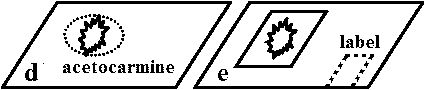 → 観察
→ 観察